
The Ediacaran Period represents that youngest part of the
Proterozoic, and is famous for the first appearance of multicellular, metazoan fossils. The Period starts at the end of the great neoProterozoic
Cryogenian, or glaciations - commonly called "Snowball Earth" - at around 635 million years ago, and ends at the start of the Cambrian (and
Phanerozoic) at around 542 million yeras ago. While a few Ediacaran stragglers may appear in the Lower Cambrian, no unambiguous multicellular/metazoan fossils have been found below the base of the Ediacaran Period, despite several possible candidates (e.g
here and
here).
In a
new paper, Meert
et al. report the possible occurrence of Ediacaran fossils from 760-770 million year old Cryogenian rocks of Kazakhstan. If correct, this would extend the fossil record of metazoans back another 100 million years. Now, there is some
evidence to suggest that metazoans were around in the Cryogenian, if only at the sponge-grade of organisation, but this is from biochemistry, not body fossils. So this would be the oldest metazoan body fossils yet found.
Two questions to be asked then. Are these deposits c. 770 million years old? Are these specimens examples of Ediacaran fossils?
I think the answer to the first question is yes, and the answer to the second question is no. I'll explain below.
AgeI have to say that Meert
et al. have done an excellent job of mapping and correlating what appears to be a very difficult sequence. It's condensed and so quite thin (by contrast, the Ediacaran type section in Australia is several kilometres thick), a significant portion missing, and the area has been subsequently affected by tectonics.

Stratigraphic column for the Lesser Karatau sequence showing the relative locations of the fossil discoveries. The fossils under discussion are from the Upper Riphean Kurgan Formation (R3) (in brown) and the glacial deposits (in yellow) and cap carbonate represents the end of the Cryogenian glacial episode and the start of the Ediacaran Period. (Meert et al. 2010)
Now with a section like this it could be argued that the glacial rocks and cap carbonate represent glacial activity known to occur in the Ediacaran and that the underlying rocks are, in fact, Ediacaran. But I think Meert
et al. have compiled enough evidence to support their stratigraphy. This includes radiometic dating of two separate samples from the Kurgan Formation that support a c. 770 million year age, as well as a δ13C stable isotopic curve that fits their interpretation (given the fragmentary nature of the curve, it could fit a number of scenarios, but it does fit this one.)
So notwithstanding the difficulties of the section, the date appears well supported. We appear to be dealing with c. 770 million year old rocks.
Fossils1) Disc and stalk
Meert
et al. have found structures in the c. 770 million year old Kergan Formation which they suggest may be referable to the Ediacaran forms
Nimbia and
Aspidella, as well as a second line of evidence, a possible stem extension emanating from the discoid fossil, and ask the question, are these Ediacaran fossils? As I said, I think the answer is no.
To show why, firstly I need to explain a bit about Ediacaran fossil preservation and positive/negative epi/hyporelief.
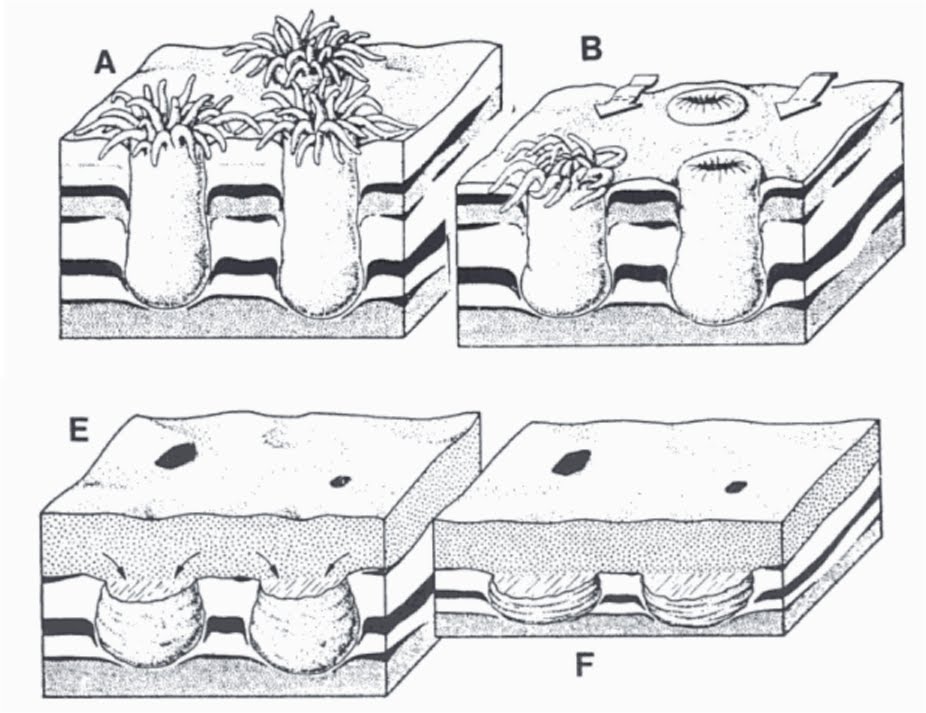
Ediacaran fossils occur primarily as marking on the surface of sandstones. These surface markings can be of two types, positive - standing up from the surface like a pimple, or negative - a depression in the surface like a dimple. Also the sandstone has two surfaces, a top surface - the epi-, and a lower surface - the hypo. Thus when we talk about surface structure, or "relief" - we talk of structure on the top bedding surface - epirelief, and structure on the lower bedding surface - hyporelief. So positive epirelief is a raised area on the top surface and negative hyporelief is a depression in the lower surface.

This is important because Ediacaran fossils occur as positive or negative relief on the bedding surfaces, but almost always as positive or negative hyporelief - that is, on the bottom surface.
The reason for this is that preservation is by sand covering a surface that has animals on it. The sand (which will eventually turn into sandstone) covers the animals. Where the animal is tough, it will push into the overlying sand, causing a negative hyporelief impression (of the top surface of the animal) into the bottom surface of the sand. Where the animal is buried in the underlying sediment, such as holdfasts, the sand flows into the depression caused by the collapse of the holdfast during burial, causing a positive hyporelief impression.

The example above is of the lower surface, The Form D projects out from the surface and so is a positive hyporelief impression (the overlying sand has pushed down into the Form D
stolon structure to form the impression. By contrast, the
Parvancorina has pushed up into the overlying sand and so is preserved as a negative hyporelief impression.
OK, lets deal with the second line of evidence first - the possible stem extension emanating from a possible discoid fossil.

(A) Discoidal impression (negative relief) with possible stem-like extension. (B) Sketch showing the location of discoidal fossil and possible stem. Meert et al. 2010.

Just to be clear about what is being discussed, here (at left) is a modern sea pen (
photo credit) we can use as an example of the frondose forms around in the Ediacaran. The structure half-buried in the sediment is the disc-shaped holdfast. Underneath the holdfast is a thin tube - the peduncle - which acts like the foot of a clam and aids in digging the organism into the sediment. The stem and frond extend out into the water column and the whole thing is held up by water pressure. The organism can inflate by pumping water into the holdfast to allow it to act as an anchor, and also into the stem and frond to hold them rigid. Pumping water out deflates the organism. Modern sea pens will deflate on touching, and collapse down onto the sediment, as a method to avoid predation or damage.
According to Meert
et al., their find may represent a holdfast and part of the attached stem that has made an impression in the sediment, in negative relief. The authors do not say if the surface is the top or bottom (and they may not know if the sample was collected loose and not found
in situ.
However, holdfast impressions are almost always found as
positive hyporelief impressions, i.e. they stick out from the lower surface. They are not found as negative relief, i.e. as depressions, as the Kurgan Formation specimen is. To find out why that is, let's look at an example.

Three-dimensional positive hyporelief fossil of a holdfast and stem from the Ediacaran of South Australia. HF impression of holdfast, IS intermediary surface within the sandstone, LS lower surface of sandstone, P peduncle, S impression of stem, W wrinkles in top surface of holdfast. White arrow is the direction of current depositing the sand.
Above is an actual Ediacaran holdfast and stem fossil. There's a lot going on here (compared with the Meert
et al. specimen where no structure is present) so it'll take a bit of explaining.
Firstly observe that this is in positive hyporelief - it's on the bottom surface of the bed. As I said, pretty much all holdfast fossils with stems are positive hyporelief. This is because the organism is sitting in the underlying sediment and gets inundated and covered by a mantling sand deposit which will become the overlying sandstone bed. Since frondose forms will deflate on contact with the enveloping sandstone, the organism pumps out water and collapses. This means that the stem falls over, usually in the direction of sand transport. The stem often gets trapped within the sand and rarely comes to rest on the sea floor because the sand has started to deposit as the organism responds. This normally results in the stem coming to rest at an shallow angle to the sea floor/base of the sand body - that is, it sticks up at an angle into the sand rather than laying down flat. This can be seen in the Ediacaran example shown above. The stem impression (
S) can be seen heading off at the ten o'clock direction but it is obviously heading off
into the sandstone - the edge of the stem impression is lower in the sandstone than the impression of the holdfast, and, as this specimen is shown upside down, the stem in heading
upwards into the overlying sandstone.
The wrinkling texture is the upper surface of the holdfast reacting to the stem being pushed over and collapsing and shows that the holdfast stayed in place during burial.
The organism is now covered by a layer of sand, with the stem trapped within the sand at a shallow angle. As the water has been pumped out, the holdfast will no longer stay inflated and so the holdfast upper surface sinks down onto the lower surface causing a shallow depression. The upper surface also takes on the shape of the lower surface where the hollow peduncle is, creating a ring shape (
P). The overlying sand then flows into the shallow depression and takes on the shape of the combined lower and upper holdfast surface structures in a process called gravity casting. Also, as the stem is angled upwards, the sand with collect behind it and be trapped between the stem and the holdfast. We can see this is the example above. The sandstone covering the stem has been removed, leaving a lighter coloured sandstone. The stem starts out on top of the holdfast, but ends up on a lower, intermediate surface (
IS) (lower from our prespective, higher when the specimen is rotated to the correct orientation).
What you end up with is a positive hyporelief impression of the combined upper and lower holdfast surface, plus the base of the stem. Furthermore, this is the only way this can be preserved. We tend not to see holdfast impressions on the tops of sandstones because the organisms appeared to favour softer, finer sediments such as silts and clays, and where active microbial mats occur.
With that in mind lets look at the Meert
et al. specimen again.

This time I've annotated it to mark out the various levels in the specimen, with
A the highest,
B an intermediary level, and
C the lowest (assuming we are looking at the top surface, or the reverse if we are looking at the lower surface).
First of all, this is a negative relief. As I explained above, holdfast and stem preservation is usually by gravity casting which produces positive relief. If we are looking at a bottom surface, maybe the organism remained intact and 'pumped up', forming a hollow in the overlying sand. But in that case the stem would also remain intact and we wouldn't see the impression of the stem in the resultant fossil. So if the stem is represented, then the organism had collapsed, in which case the holdfast would have collapsed and we would get a positive hyporelief as notmal.
The structure can't be a positive hyporelief that has eroded out the produce the depression because the original fossil is just a surface impression. The sandstone underneath the fossil is the same sandstone the rest of the rock and so there can be no preferential weathering to produce the hollow.
If we are looking at a top surface, then it is unlikely to be a holdfast and stem as we do not find them on top surfaces of sandstones. Even if it were, the stem appears to have been pushed into the sand body to a level equal to, or even greater than, the depth of the holdfast. Again an unlikely occurrence, as a slight impression on the underlying sand is the most you are likely to get.
Given that this is a negative relief and the lack of any distinguishing structure, it is most likely that the specimen is simply an erosional feature and not a fossil.
2)
Nimbia and
AspidellaSpecimens referable to
Nimbia and
Aspidella are reported from the c. 770 millio year old Kurgan Formation.
Our specimens were discovered in a brown-red shale within the largely siliclastic Kurgan Formation and are preserved in both hypo and epi-relief. They are circular to oval-shaped impressions that surround either a smooth interior or circular to oval indentations with a smooth interior. (Meert et al. 2010.)

(A) closeup of the Nimbia fossils in positive epirelief, AT marks the occurrence of a possible invaginate morph of Aspidella terranovica; (B) B&W photo of Nimbia impressions shown mostly in positive epirelief; (C) Nimbia fossils in negative relief with a possible invaginate morph of Aspidella terranovica in positive relief; (D) large c. 2 cm (long axis) c. 1.5 cm (short axis) negative hyporelief impression of Nimbia occlusa and an ‘inversion’ of the photo the fossil might appear in positive epirelief. White arrow points to a possible raised central nodule observed in Nimbia occlusa fossils elsewhere. Meert et al. 2010.
Taking
Aspidella first. Actually, attributing the form to
Aspidella doesn't tell us much. This is because
Aspidella should now be considered a form genus - that is a grouping of similar forms (i.e. discs) and not representative of a single organism, or related group of organisms. In other words, a lot of different things make disc impressions, e.g. algae (algal biscuits), microbial colonies, fungae and metazoans. Calling the specimen
Aspidella simply says that it is a certain kind of disc, not that it is the Ediacaran form
Aspidella terranovica. There are a lot of morphologies within the
Aspidella group and they are not produced by the same organism, or even closely related organisms. This means that more than one organism is included under
Aspidella. Even worse, no consistent distinction can be made between the various disk structures, making it difficult to accurately assign a particular disc to a particular organism. Where other evidence exists, i.e. stem impressions, ornamentation, etc, it is possible to identify individual organisms or groups of organisms. But simple discs remain probematic. Being a disc does not make it a holdfast!
It is entirely likely that
Aspidella actually represents a group of diverse, not closely related, organisms - microbial, algal, fungal and metazoan - that produce discs and overlap each other to give
Aspidella a large time range, but this time range is not for a single species or even genus.
This boils down to the fact that
Aspidella with stems or other ornamentation, and associated with other Ediacaran forms, can be considered a metazoan fossil. Other
Aspidella with no other ornamentation, and not associated with Ediacaran forms, could be anything.
The
Nimbia find is in a similar situation. Do we have
Nimbia? Yes. But here also, I think that the form
Nimbia, like
Aspidella, may hide a multitude of sins, or at the very least a number of different organisms. Except I think we have proof here that
Nimbia is not an Ediacaran metazoan - or at least
this Nimbia is not an Ediacaran metazoan.
The specimens of
Nimbia from the Kurgan Formation are found as positive epirelief, negative epirelief, and negative hyporelief. This does not happen for Ediacaran fossils, as discussed above. Since these
Nimbia show a number of modes of preservation, and they all show the same structure regardless of mode of preservation, it is apparent that the form that produced it is rigid. It has to be if it produced the same shape whether on the top surface creating a positive or negative imprints, or on the bottom surface. The fossil is the same independent of preservational style, and so whatever is causing the fossil must therefore be rigid like a coin. That rigidity, and the various preservational styles, means that it cannot be a holdfast.
It has been suggested that
Nimbia represents microbial colonies. Hard, well agglutinated disk-shaped colonies would be tough enough to cause the impressions, and some have been known to develop a central nodule similar to that found in some
Nimbia. This would explain the similar shape through different modes of preservation, and the large time range of the form.
Finally, Ediacaran fossils are normally found as assemblages, with a number of different forms present. The Kurgan Formation specimens are of one main type,
Nimbia, and a few
Aspidella. Too restricted to be considered an assemblage (a random collection would be expected to uncover more than two forms if an assemblage was present). Even if this find represented an early stage in the evolution of metazoa, it would be expected that more than two forms would be present.
So, discoial fossil with stem? - no.
Aspidella? - yes, possibly, but that doesn't mean it's an Ediacaran or even metazoan.
Nimbia? - yes, but probably not metazoan.
In properly posing questions about their finds instead of making pronouncements, and suggesting a number of possible interpretations for their finds, Meert at al. have highlighted the important implication of their finds.
It is possible that our discovery of Nimbia occlusa and Aspidella terranovica(?) in sedimentary rocks during the early Cryogenian (N766 Ma) lends support the alternative hypotheses regarding these fossils and remove them from consideration as true metazoa(Meert et al. 2010.)
I think they are right, they have provided evidence that the forms are not metazoan - or at least not "Ediacaran" (as in found amongst Ediacaran assemblages) - fossils, and that the simple disc-shaped forms found in the Proterozoic comprise a form group with contributions from a number of metazoan and non-metazoan sources.
Meert, J., Gibsher, A., Levashova, N., Grice, W., Kamenov, G., & Ryabinin, A. (2010). Glaciation and ~770Ma Ediacara (?) Fossils from the Lesser Karatau Microcontinent, Kazakhstan Gondwana Research DOI: 10.1016/j.gr.2010.11.008Further reading on Ediacaran discs
Gehling, J.G., Narbonne, G.M., and Anderson, M.M (2000) The first named ediacaran body fossil Aspidella terranovica. Palaeontology, 43: 427-456. DOI: 10.1111/j.0031-0239.2000.00134.xMacGabhann, B.A. (2007) Discoidal fossils of the Ediacaran biota: a review of current understanding. Geological Society, London, Special Publication, 286: 297-313. DOI: 10.1144/SP286.21














































{kind=link}
{kind=link}