
Sshhh, be vewy vewy quiet. We’re hunting Awchean pwokawyotes.
There is good carbon and sulphur isotope geochemical evidence that life has existed on Earth from the early Archean. This life is thought to consist of small prokaryote cells with a penchant for sulphur instead of oxygen.
However, identifying actual fossil evidence is very tricky for a number of reasons. One of the major ones being, that given the 3.5 billion year time span and plate tectonics, few rocks from this period of Earth history remain on the surface. Of those that do, most either were unsuitable to preserve life in the first place (e.g. volcanic rocks) or have been metamorphosed until any possible organic trace has been cooked to oblivion.
So, if you want to go hunting Archean microbes you need to look in an area with rocks that have been relatively unmolested for some 3.5 billion years or so.

This is the Strelley Pool Formation and is thought to be between 3426-3340 million years old. At the base of Formation, sitting unconformably on underlying 3515 million year old basalts, is a remarkably well preserved sandstone. (The ridge in the header image above. The red dots mark the position of the unconformity between the underlying basalts and the Strelley Pool Formation. Image (sans Elmer) from Wacey et al. 2011. Supplementary Information)
The sandstone is dark coloured at the base, due to it being carbonaceous- and pyrite-rich, where it sometimes fills channels eroded into the underlying basalt. Above this, the sandstone becomes lighter as the carbonaceous matter and pyrite disappear, before passing into overlying limestones - which contain possible stromatolites - and cherts.
 |
| Thin section photomicrograph showing multiple generations of carbonaceous material and silica cements within the Strelley Pool sandstone. DQ, detrital quartz grains. C1, carbon material, interpreted as a biolfilm, covering quartz grains. S1, first silica cement layer, up to c. 30 μm partially filling the void space between the detrital quartz grains. C2, A second carbon biofilm covering the surface of the S1 silica layer. S2, a second generation of silica cement precipitated in the larger pore spaces. C3, a third generation of carbon biofilm. (Wacey et al. 2011. Supplementary Information). |
A second film of carbonaceous matter covers the primary silica cement layers. The central spaces between grains not occupied by the silica layers and carbon films, is filled with chert.
From this we can say that the well-rounded, large quatrz and pyrite grains suggest the sandstone was deposited in a high energy environment, such as a beach or estuary. The sandstone is also very clean - there is no fine material such as silt or clay - also supporting a high energy environment of deposition which would have washed out all the fine-grained material such as silt and clay.
The several episodes of silica cementation suggests cyclic inundation of silica-rich fluids into porous, clean sands.
 |
| Thin section showing b, Rounded quartz grain separated from a carbon-rich chert infilling by a dripstone cement (white arrow). c, Rounded grain of detrital pyrite (black grain) with a dripstone chert cement and carbon coating (red arrow). (Wacey et al. 2011). |
The evidence suggests then, that the sandstone was deposited in a nearshore beach-type environment which helped smooth out the quatrz and pyrite grains, and removed any fine material. During this time the first layer of carbonaceous material was formed around the grains (C1 above). Periodically, the highly porous sand came under the influence of valdose conditions, allowing the precipitation of silica cement around the grains (S1 above), trapping the carbonaceous layer, but maintaining the porous nature of the sand. Subsequently a second layer of carbonaceous material (C2 above) was formed over the S1 cement, possibly suggestive of a reversion to marine influences. A second round of valdose conditions produced a second cementation layer (S2 above). Followed by another return to marine influences and a third carbonaceous layer (C3 above). Finally, after the sands were buried, subsequent chert deposition provided the source for the last event, the filling of the remaining pore space with chert.
Before we look at the carbonaceous material in more detail there is the question of the link between the detrital pyrite grains and the carbonaceous matter.
The detrital pyrite grains are large, much larger than the possible cells in the carbonaceous matter, and are rounded. This suggests that the pyrite is not a product of possible organic activity. There is some pyrite within the carbonaceous matter, but this pyrite is much smaller and angular in shape - as expected if they have been precipitated as a result of organic activity. The detrital pyrite grains are also covered by the silica cement layers. The evidence suggests that the pyrite is primary - that is it was present as an original constituent of the sand - rather then being a product of organic activity. As the pyrite only occurs in the base of the sandstone, primarily in erosive channels in the underlying volcanics, it seems likely that the pyrite grains were formed from the erosion of underlying rocks and have been left behind as a lag deposit as pyrite grains are much heavier that similar sized grains formed from other materials like quartz. The renewed influx of sediment resulted in the pyrite grains mixing with the quartz grains to form a pyrite-rich basal sand. Subsequent deposts of sand did not have the pyrite grains and so were deposited free of pyrite and carbonaceous matter. Remember also that the 3.4 Bya atmosphere has precious little in the way of free oxygen, so the pyrite would be quite stable under surface conditions.
Since the detrital pyrite and carbonaceous matter are absent from the sandstone above the basal layer, there does appear to be a connection between the detrital pyrite and the carbonaceous matter. It appears that the carbonaceous matter is present because of the pyrite.
To answer why the carbonaceous matter should be linked to the pyrite, we first need to know what the carbonaceous matter is and what it contains.
 |
Ramen spectroscopy (it's to do with lasers and nothing to do with Arthur C. Clarke) supports a disordered structure. This is important because organic matter is disordered, inorganic carbonaceous matter is ordered - abiotic graphite, for example, is found in rocks of this age but shows a well ordered internal structure. So that's a tick for organic. Also, an analysis of the structures indicates the presence of carbon, nitrogen and sulphur in the possible cell wall structures. Again a tick for organic as these are essential for organically formed structures, but not for inorganic ones.. Finally analysis of the carbon in the possible cell walls shows a 26-39 parts per thousand enrichment in 12C compared with atmospheric CO2. This also support the structures being organic as only organic activity can partition light 12C from the 'heavier' 13C to such an extent.
So the evidence suggests that the structures are organic, but are they microfossils?
Well, the shapes are round and tube-shaped. So, as prokaryotes tend to be round and tube-shaped, this is a good thing. However, a number of inorganic processes can produce round and tube shapes. So, by itself, this doesn’t tell us much. The specimens do show some evidence of folding, which organic shapes would do that inorganically produced shapes wouldn’t. So shape is a point in favour of them being microfossils, but not conclusive.
Size is an interesting one. Prokaryotes come in a range of sizes, but modern forms are generally around the 10 μm mark. The forms found range in size from 5-25 μm, with an average size of around 10 μm, not a bad comparison with modern forms.
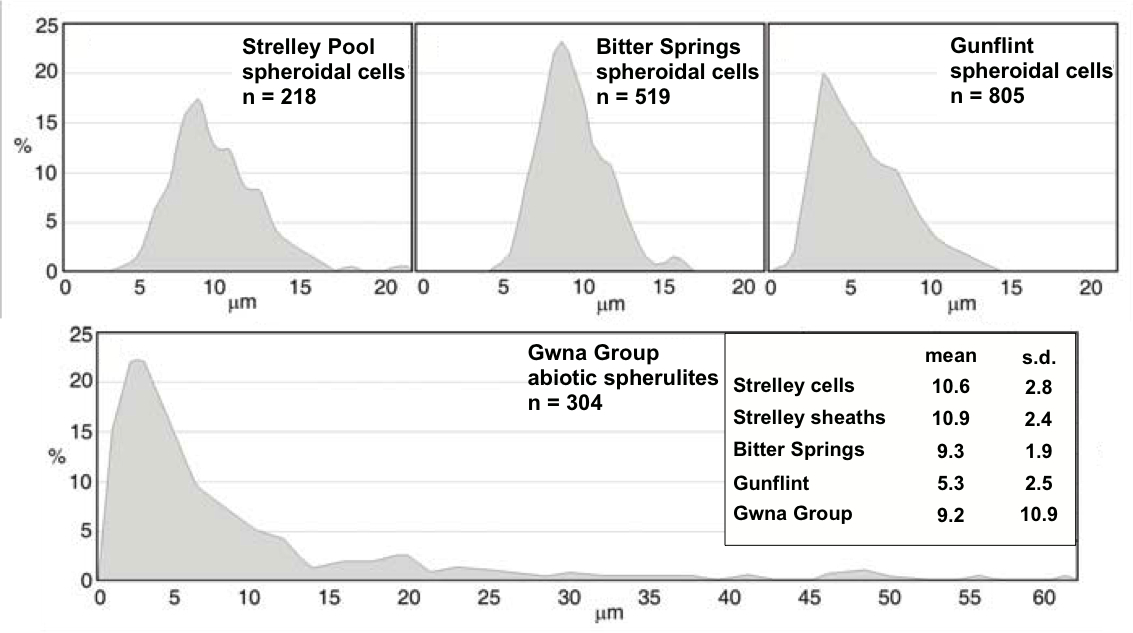 |
| Size distributions of microfossils from the Strelley Pool Sandstone compared to those from three younger biological microfossil assemblages and one abiotic spherulite assemblage. (Wacey et al. 2011). |
 |
| 80 μm giant from Wacey et al. 2011 |
Another explanation could be that these large ‘cells’ represent a cluster of cells in which only the outer cell walls in contact with pore fluids have been preserved, while the internal walls dividing individual cells are not preserved. Certainly the structures arrowd in the large specimen could represent the boundaries between cells as much as they could represent folding. The presence of silica micro-crystals in cell walls does suggest that silica-rich pore fluids played a part in the preservation process. So the larger specimens could be the remains of clumps of cells, or possibly a different group of organisms from the smaller ones.
The numerous lines of evidence tends to support the identification of these structures as organic ans potentially prokaryote cells, making them the oldest life forms so far identified.
So why the attraction to sulphur? Well, in the absence of oxygen, SO42- makes a good terminal electron acceptor. Larry Moran has the details.
Given the age, it will probably be impossible to get absolute evidence that any particular set of structures represent Archaen prokaryotes, but the evidence Wacey et al. 2011 present is probably as close as we could reasonably expect to come.
So with these Archaen prokaryotes, and the Ediacaran fauna, when it comes to Precambrian life, AUSTRALIA ROCKS!
Wacey, D., Kilburn1, M.R., Saunders, M., Cliff, J. and Brasier, M.D (2011) Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia. Nature Geoscience. Published online Aug. 21. DOI: 10.1038/NGEO1238



