Things have been a bit quiet recently because inexplicably, the people who pay me actually want me to do stuff!
Part of that stuff will have me at a loose end in Hamburg on the weekend of 11th and 12th June. So if anyone has any suggestions about things to do in Hamburg, I'd be grateful for some tips.
Meanwhile, normal service will be resumed as soon as possible
Friday, May 20, 2011
Sunday, April 3, 2011
Mopping up some Ediacaran Enigmatics

{kind=link}
A good example of this are "mops".
"Mops" are a series of abundant, distinct structures which look like the head of a mop - hence the name. "Mops" are always preserved in hyporelief, that is on the underside of the rock, unusually though, they can be either positive - standing up from the surface like a pimple, or negative - a depression in the surface like a dimple. Most Ediacaran fossils are usually either one or the other.
Tarhen et al. studied a series of "mops" which show a highly varied shape and structure, but found some consistent features.

Scale bar = 2cm. (Tarhan et al. 2010)
(1) "Mops" are always orientated within 10 degrees of the palaeocurrent as derived from a number lines of evidence.
(2) They have a distinct margin at one end - considered to be the distal or far end. This can be straight (arrowed in (A) above), curved (in (B)) or lumpy (in (C)).
(3) They have a series of roughly parallel lines running at around 90 degrees to the margin (arrowed in (B) above). These appear filament-like and can be either linear or wavy and tangled
These features can vary even within the same bed and between specimens in close proximity.
They do tend to appear in close association with the disc form Aspidella, which it though to represent the holdfast of a Charniodiscus-like organism or sea pen. But are they fossils or non-biogenic marks?
Well they appear biogenic. The association with Aspidella is too strong to be random. But are they body or trace fossil? Well, they are a little of both.
The association with Aspidella appears important. Aspidella is the holdfast of a sea pen-like organism, which had a large bulbous holdfast buried in the sediment, with a stalk and frond rising up into the water column. So the organism has some elements (the holdfast) below the bacterial mat, and some (the frond) above the mat.
In this configuration, the frond will be subject to water currents. What is though to have happened is that strong currents carrying the sand that will eventually overlay the organisms has hit the frond and basically dragged the whole organism including the holdfast, in the direction of the palaeocurrent. As the holdfast is under the mats, this dragging has uprooted the holdfast and dragged it through the microbially bound mat layer. The parallel lines represent torn-up bits of mat which were attached to the top of the holdfast. Lumpy margins of the 'mops' are probably caused by small lumps of sediment trapped next to the holdfast.

D: Sand deposition. E: Preservation. (Tarhan et al. 2010)
This explanation can explain a number of structures seen in Ediacaran rocks. One in particular is Pseudorhizostomites.

Pseudorhizostomites has been interpreted as a rhizostomean medusa (a jellyfish without tentacles), and even a gas escape structure formed during the decay of an organism. But an interesting interpretation based on the 'mop' formation, is that they are Aspidella-like holdfasts that have been torn free by current action, but vertically rather than in a more horizontal direction. So they have been plucked out of the sediment rather than being dragged along.
Another structure that appears related is one associated with actual holdfasts.

Here the holdfast has remained intact and in place, but the stem has been flattened and the surface of the holdfast has been distorted by the sheer forces caused by the current pushing the frond and stem over.

Here is an annotated example that has been featured here before. The wrinkles (W) and the impression of the stem (S) are clearly visible and have been caused by the stem and frond being pushed over by a strong current which eventually deposited the overlying sand bed
The explanation of the "mop" structures is a neat use of the interplay between sedimentology and taphonomy, that ties together a number of structures.
Tarhan, L.G., Droser, M.L. and Gelhing, J.G (2010) Taphonomic Controls on Ediacaran Diversity: Uncovering the Holdfast Origin of Morphologically Variable Enigmatic Structures. Palaios V. 25, pp 823-830. DOI:10.2110/palo.2010.p10-074r
Friday, April 1, 2011
New Find Challenges Evolution
I've been a passionate supporter of evolution, but now . . .
A recent find from the Flinders Ranges of South Australia will shake the theory of evolution to the core. But no-one knows about it.
Why?
I've tried to talk to palaeontologists about it, but the refuse to discuss it. Evolutionists I've known for years no longer reply to my emails. I've been ignored buy organising committees for conferences and symposiums in favour of other evolutionists when I have earth shattering evidence against their pet theory.
They may have frozen me out, but I will not be silenced.
From a Precambrian site deep in the Flinders Ranges, I was guides by something . . . or someone, and I found startling remains - reptile remains with clear preservation of skin. How could that be if the 'so called' (in my opinion now) theory of evolution is true? How can exceptionally preserved reptile remains be present at a Precambrian site?
It can't. But it is. So evolution must be false.
Click on the link below and spread the evidence the the evolutionists have tried to stop me from exposing.
Reptiles in the Precambrian disproves evolution

A recent find from the Flinders Ranges of South Australia will shake the theory of evolution to the core. But no-one knows about it.
Why?
I've tried to talk to palaeontologists about it, but the refuse to discuss it. Evolutionists I've known for years no longer reply to my emails. I've been ignored buy organising committees for conferences and symposiums in favour of other evolutionists when I have earth shattering evidence against their pet theory.
They may have frozen me out, but I will not be silenced.
From a Precambrian site deep in the Flinders Ranges, I was guides by something . . . or someone, and I found startling remains - reptile remains with clear preservation of skin. How could that be if the 'so called' (in my opinion now) theory of evolution is true? How can exceptionally preserved reptile remains be present at a Precambrian site?
It can't. But it is. So evolution must be false.
Click on the link below and spread the evidence the the evolutionists have tried to stop me from exposing.
Reptiles in the Precambrian disproves evolution
Thursday, February 17, 2011
Paris Feb 2011
Something told me I'd picked the wrong day to check out the view from the top of the Eiffel Tower . . .




Wednesday, February 2, 2011
Palaeoporn 22

The type section for the Cambrian-Precambrian boundary is at Fortune Head, Burin Peninsula, Newfoundland, as is marked by the first appearance of the trace fossil Treptichnus pedum (the trace fossil formally known as Phycoides pedum).
Treptichnus pedum is a distinctive burrow pattern comprised of a series of lobes set along a central, sometimes curved burrow, thought to represent successive probes into the sediment searching for food. The traces are thought to represent an organism more complex that those of the Ediacaran, but that is not necessarily true, as no actual animal has been found is association with the burrows, so we don't know what made them (although modern priapulid worms make similar burrows).
However, we should not confuse the somewhat complicated pattern of Treptichnus pedum with the first occurrence of well-developed, fairly complex metazoan animals.
While the start of the Cambrian is marked by the first appearance of Treptichnus pedum, this is not the first appearance of trace fossils probably from metazoans. A number of trace fossil types exist in the preceding Ediacaran rocks. In fact they show a neat line in increasing complexity from simple traces early in the Ediacaran to traces as complex as T. pedum towards the end of the Ediacaran.
The earliest traces we have, and the most simple, is Planolites

This is a lowermost Cambrian specimen (picture from NCSE), but it shows you what they are like. The fossil is of simple, cylindrical, unbranched and unlined burrows. They are usually sinuous and undulatory, and often appear as small knobs or discontinuous segments on bedding planes. The small image below is what they typically look like in Ediacaran rocks

Further up in the Ediacaran, more traces appear. This is Helminthopsis

Helminthopsis are much more meandering burrows that appear to represent shallow feeding burrows. the distinctive whorls and loops are considered to represent a different feeding strategy to that of Planolites.
Towards the top of the Ediacaran, things get a bit more complex. As well as Planolites and Helminthopsis, we get the feeding traces of Kimberella.

Here, the feeding traces are marked "R" for radula, and the animal itself is marked "K". A Dickinsonia is also present. "K'" and "D'" mark the resting trace of Kimberella and Dickinsonia, which comprise, of course, another form of trace fossil.
Towards the top of the Ediacaran, things start to get crowded. Not only do we have all the previous traces, but they are joined by Mattaia miettensis.

This specimen comes from the Kessyusa Formation, from Khorbusuonka in northern Siberia. The trace is filled with two bands of sediment, raised on the sides and depressed in the middle, where the bands are separated by a vertical fissure filled with clay. A possible culprit is a pripulid worm such as the Burgess Shale form Louisella pedunculata.
So by the time you get to the Cambrian-Precambrian boundary there were quite a few traces around. Sure in the Cambrian the traces get much more diverse, but they don't start at the boundary, there is a steady increase in trace fossils through the Ediacaran.
Update
Aleksey Nagovitsyn has kindly informed me that the age range for Mattaia miettensis in Palaoeporn 22 is incorrect, and Mattaia miettensis is actually to be found in the Cambrian Tommotian Stage of the Cambrian.
So we need to remove Mattaia miettensis from the list of trace fossils found at the top of the Ediacaran.
Friday, January 14, 2011
"You Cannot Trust Reasons to Believe"
At least that's what Todd Wood thinks.
In a series of detailed posts, Todd does an excellent job of taking apart a series of posts and responses by Dr Fuzale Rana of Reasons to Believe (a christian group that "bridges the gap between science and faith by exploring questions about God and the Bible". They have no problem with an ancient Earth, but evolution is apparently another matter) on the similarity between the human and chimp genome (or not, in the case of Reasons to Believe) and moves on to pseudogenes, thus:
Well, you may say, 'showing up the mangling of science by those seeking to support their religious views, is not unusual".
True. But Todd happens to be a young earth creationist.
Double ouch!
Here is post one, from there move on to "Newer Post" and read them in sequence. Post four is at the bottom of the front page of Todd's blog. You can work your way up.
It's well worth the read.
In a series of detailed posts, Todd does an excellent job of taking apart a series of posts and responses by Dr Fuzale Rana of Reasons to Believe (a christian group that "bridges the gap between science and faith by exploring questions about God and the Bible". They have no problem with an ancient Earth, but evolution is apparently another matter) on the similarity between the human and chimp genome (or not, in the case of Reasons to Believe) and moves on to pseudogenes, thus:
"In his latest post, Rana asserts the following about the argument for common ancestry from pseudogenes:Ouch!
When evolutionary biologists present this argument, they make a number of assumptions, all of which appear to have questionable validity based on recent research results. For the pseudogene evidence to have potency: (1) pseudogenes must lack function; (2) their origin must be due to rare, random events; and (3) their juxtaposition to other genes must be arbitrary.
Everything he wrote there is utterly false. None of those conditions are required to argue for common ancestry from pseudogene similarity. Not one."
Well, you may say, 'showing up the mangling of science by those seeking to support their religious views, is not unusual".
True. But Todd happens to be a young earth creationist.
Double ouch!
Here is post one, from there move on to "Newer Post" and read them in sequence. Post four is at the bottom of the front page of Todd's blog. You can work your way up.
It's well worth the read.
Sunday, January 9, 2011
Palaeoporn 21

In keeping with the positive hyporeliefian, is-it-or-isn't-it-a-metazoan theme of the last couple of posts, here is Beltanelliformis brunsae. A widespread form (this one is from the Mackenzie Mountains of Canada) that has had a history of being moved around the tree of life.
Notice firstly the texture of the surface. This is a microbial mat that has undergone some considerable deformation, probably due to the sudden influx of sand which pressed down on the mat and caused it react unevenly to the sudden load.
The Beltanelliformis appear at first glance to be similar to other disc-like Ediacaran forms associated with holdfasts. However, notice that the specimens pictures are in strong positive hyporelief, that is they extend quite some way out from the lower surface of the sandstone. This means that the sand infilled quite a sizable hole in the underlying mud. Notice also that, in the top two specimens, the fossil has a 'collar' around the basal disc, which makes the hole infilled by the sand cone-shaped, with the base being the smaller diameter part of the cone.
This configuration let to suggestions that the form represents the burrow of an anemone.
Below is a representation of that interpretation (from Schopf et al.).
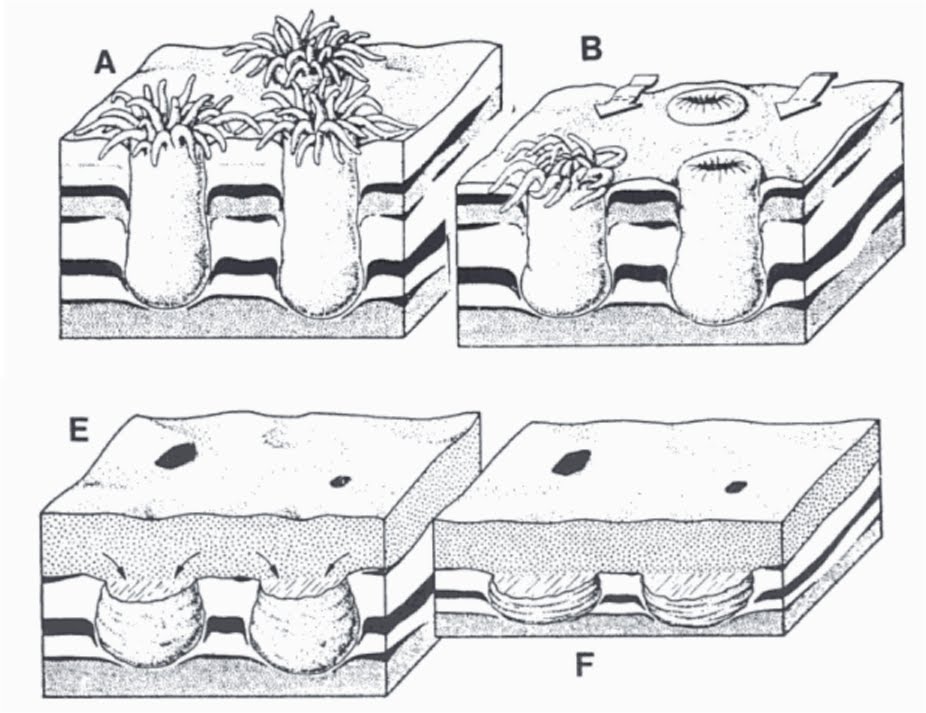 However, more recent interpretations have Beltanelliformis as an algae.
However, more recent interpretations have Beltanelliformis as an algae.What!? Algae!?
Yes, algae. Here's why.
The uppermost Doushantuo Formation (590-555 Ma) at Miaohe in the Yangtze Gorges area, is a series of dolomites that provide a rare Burgess-Shale-type taphonomic window on the Ediacaran. Within the Formation are found numerous carbonate compressions of organisms. This represents a different mode of preservation to that of the more typical sandstone preservation, at Miaho the forms are flattened carbonate compressions. Amongst the forms present are several that are interpreted as Beltanelliformis

Beltanelliformis brunsae from the Doushantuo Formation. Scale in 2, 1 cm. from Xiao et al. 2002)The non-overlapping nature of the fossils is a good indication that we are dealing with benthic (bottom-dwelling) forms and not free-swimming forms that have died and sunk to the bottom. It may well be that the forms were actually tethered to bottom with a small holdfast structure.
The Doushantuo specimens have thing flexible walls. However, given the original specimen at the beginning of this post, it's clear that they would need to retain the three-dimensional shape during burial by sand.
A modern form that appears to fit the bill is the caulerpalean green alga Derbesia (photo credit).

Derbesia is benthic, is attached by a minute holdfast, and has the right shape and size. Also, it is organized as a coenocytic film of cytoplasm within a elastic and strong wall and surrounding one or more large, fluid-filled vacuoles. These vacuoles would be sealed and so couldn't empty on burial like holdfasts could. This makes it strong enough to make an impression in sandstone preservation!
So Beltanelliformis has gone from Cnidarian -> holdfast -> anemone burrow -> algae with holdfast.
Interestingly, apart from Nimbia and Aspidella mentioned in a previous post, Beltanelliformis has one of the longest time ranges, having been found in the pre-Ediacaran Cryogenian in Canada.
This again raised the possibility that several of the generic Ediacaran disc fossils could be something other than metazoans. That's actually fine. We have enough evidence that some of the discs represent metazoans. That some don't, should be expected. After all, if the Ediacaran assemblages represent vibrant, healthy ecosystems, they would be expected to contain microbial, algal, and metazoan, life.
Monday, January 3, 2011
To Epirelief or Hyporelief, That is the Question . . .
My last post on the possible 770 million year old Ediacaran fossils from Kazakhstan prompted some comments regarding epi- and hyporelief (go read the post and comments).
Basically, Ediacaran fossils are usually found on the underside of the sandstone bed in either positive relief (sticking out from the surface) or negative (sunk into the surface) hyporelief (hypo- is sciencey for 'on the underside'). This is dictated by the mode of preservation (again see the original post), and means that we do not find them on the top surface of the sandstone bed, or in epirelief. (Actually we do find them in epirelief, but only at Mistaken Point in Newfoundland where they have been mantled by volcanic ash - but that appears to be a one off and so does not represent normal preservation).
As an example here is a slab I collected from the Flinders Ranges in South Australia. First the upper (epi) surface.

As you can see, its rippled - strongly rippled actually - which puts pay to the ideas that the Ediacaran faunas were deep water, but that's another story. However, what the top surface isn't, is fossiliferous. There are no fossils on it.
Now lets take a look at what's underneath.

Ta Da! This was taken in situ, about 5 minutes after I'd found it. The photo is a bit blurry 'cos my hands were stili shaking!
Notice the smooth lower surface. This is because it is mantling a mud which does not form ripples. Also notice that there are three large discs in positive hyporelief and a Dickinsonia in negative hyporelief. What do you mean no!?. Ok, here's an annotated version.

The question has been asked why the counterpart of a positive hyporelief isn't a negative epirelief on the bed below. In other words if the fossil is formed by sand flowing into a depression in the underlying mud to form a positive hyporelief, should there also be a corresponding negative epirelief on the top of the underlying bed?
The answer is yes, but we almost never find them. The reason is that the underlying bed is made of mud and so compresses during diagenesis which disrupts the fossil. However, the main reason is that the mudstone weathers much more readily than the sandstone and forms a very friable, crumbly rock that simply weathers away.
We do have some examples of underlying epirelief fossils, but they are very rare. To find them you have to excavate the sandstone with the underlying mudstone still in place. Even then, they are very fragile. Normally this never happens in nature as the mudstone erodes away. The fossiliferous sandstone then weathers out and falls downslope to be found by palaeontologists.
Below is a typical Ediacaran location. The fossils are eroding off the top of the hill and can just be seen in outcrop. but the fossils are found on the scree slope, as float - that is, material that has eroded out of the outcrop and has slid downslope

So the answer is, yes there are there epirelief fossils but we almost never find them as they have weathered away. But they would be in mudstones and not in sandstones.
Basically, Ediacaran fossils are usually found on the underside of the sandstone bed in either positive relief (sticking out from the surface) or negative (sunk into the surface) hyporelief (hypo- is sciencey for 'on the underside'). This is dictated by the mode of preservation (again see the original post), and means that we do not find them on the top surface of the sandstone bed, or in epirelief. (Actually we do find them in epirelief, but only at Mistaken Point in Newfoundland where they have been mantled by volcanic ash - but that appears to be a one off and so does not represent normal preservation).
As an example here is a slab I collected from the Flinders Ranges in South Australia. First the upper (epi) surface.

As you can see, its rippled - strongly rippled actually - which puts pay to the ideas that the Ediacaran faunas were deep water, but that's another story. However, what the top surface isn't, is fossiliferous. There are no fossils on it.
Now lets take a look at what's underneath.

Ta Da! This was taken in situ, about 5 minutes after I'd found it. The photo is a bit blurry 'cos my hands were stili shaking!
Notice the smooth lower surface. This is because it is mantling a mud which does not form ripples. Also notice that there are three large discs in positive hyporelief and a Dickinsonia in negative hyporelief. What do you mean no!?. Ok, here's an annotated version.

The question has been asked why the counterpart of a positive hyporelief isn't a negative epirelief on the bed below. In other words if the fossil is formed by sand flowing into a depression in the underlying mud to form a positive hyporelief, should there also be a corresponding negative epirelief on the top of the underlying bed?
The answer is yes, but we almost never find them. The reason is that the underlying bed is made of mud and so compresses during diagenesis which disrupts the fossil. However, the main reason is that the mudstone weathers much more readily than the sandstone and forms a very friable, crumbly rock that simply weathers away.
We do have some examples of underlying epirelief fossils, but they are very rare. To find them you have to excavate the sandstone with the underlying mudstone still in place. Even then, they are very fragile. Normally this never happens in nature as the mudstone erodes away. The fossiliferous sandstone then weathers out and falls downslope to be found by palaeontologists.
Below is a typical Ediacaran location. The fossils are eroding off the top of the hill and can just be seen in outcrop. but the fossils are found on the scree slope, as float - that is, material that has eroded out of the outcrop and has slid downslope

So the answer is, yes there are there epirelief fossils but we almost never find them as they have weathered away. But they would be in mudstones and not in sandstones.
Sunday, January 2, 2011
770Ma Ediacara (?) Fossils from Kazakhstan (sadly no)
 The Ediacaran Period represents that youngest part of the Proterozoic, and is famous for the first appearance of multicellular, metazoan fossils. The Period starts at the end of the great neoProterozoic Cryogenian, or glaciations - commonly called "Snowball Earth" - at around 635 million years ago, and ends at the start of the Cambrian (and Phanerozoic) at around 542 million yeras ago. While a few Ediacaran stragglers may appear in the Lower Cambrian, no unambiguous multicellular/metazoan fossils have been found below the base of the Ediacaran Period, despite several possible candidates (e.g here and here).
The Ediacaran Period represents that youngest part of the Proterozoic, and is famous for the first appearance of multicellular, metazoan fossils. The Period starts at the end of the great neoProterozoic Cryogenian, or glaciations - commonly called "Snowball Earth" - at around 635 million years ago, and ends at the start of the Cambrian (and Phanerozoic) at around 542 million yeras ago. While a few Ediacaran stragglers may appear in the Lower Cambrian, no unambiguous multicellular/metazoan fossils have been found below the base of the Ediacaran Period, despite several possible candidates (e.g here and here).In a new paper, Meert et al. report the possible occurrence of Ediacaran fossils from 760-770 million year old Cryogenian rocks of Kazakhstan. If correct, this would extend the fossil record of metazoans back another 100 million years. Now, there is some evidence to suggest that metazoans were around in the Cryogenian, if only at the sponge-grade of organisation, but this is from biochemistry, not body fossils. So this would be the oldest metazoan body fossils yet found.
Two questions to be asked then. Are these deposits c. 770 million years old? Are these specimens examples of Ediacaran fossils?
I think the answer to the first question is yes, and the answer to the second question is no. I'll explain below.
Age
I have to say that Meert et al. have done an excellent job of mapping and correlating what appears to be a very difficult sequence. It's condensed and so quite thin (by contrast, the Ediacaran type section in Australia is several kilometres thick), a significant portion missing, and the area has been subsequently affected by tectonics.

Stratigraphic column for the Lesser Karatau sequence showing the relative locations of the fossil discoveries. The fossils under discussion are from the Upper Riphean Kurgan Formation (R3) (in brown) and the glacial deposits (in yellow) and cap carbonate represents the end of the Cryogenian glacial episode and the start of the Ediacaran Period. (Meert et al. 2010)Now with a section like this it could be argued that the glacial rocks and cap carbonate represent glacial activity known to occur in the Ediacaran and that the underlying rocks are, in fact, Ediacaran. But I think Meert et al. have compiled enough evidence to support their stratigraphy. This includes radiometic dating of two separate samples from the Kurgan Formation that support a c. 770 million year age, as well as a δ13C stable isotopic curve that fits their interpretation (given the fragmentary nature of the curve, it could fit a number of scenarios, but it does fit this one.)
So notwithstanding the difficulties of the section, the date appears well supported. We appear to be dealing with c. 770 million year old rocks.
Fossils
1) Disc and stalk
Meert et al. have found structures in the c. 770 million year old Kergan Formation which they suggest may be referable to the Ediacaran forms Nimbia and Aspidella, as well as a second line of evidence, a possible stem extension emanating from the discoid fossil, and ask the question, are these Ediacaran fossils? As I said, I think the answer is no.
To show why, firstly I need to explain a bit about Ediacaran fossil preservation and positive/negative epi/hyporelief.
Ediacaran fossils occur primarily as marking on the surface of sandstones. These surface markings can be of two types, positive - standing up from the surface like a pimple, or negative - a depression in the surface like a dimple. Also the sandstone has two surfaces, a top surface - the epi-, and a lower surface - the hypo. Thus when we talk about surface structure, or "relief" - we talk of structure on the top bedding surface - epirelief, and structure on the lower bedding surface - hyporelief. So positive epirelief is a raised area on the top surface and negative hyporelief is a depression in the lower surface.
 This is important because Ediacaran fossils occur as positive or negative relief on the bedding surfaces, but almost always as positive or negative hyporelief - that is, on the bottom surface.
This is important because Ediacaran fossils occur as positive or negative relief on the bedding surfaces, but almost always as positive or negative hyporelief - that is, on the bottom surface.The reason for this is that preservation is by sand covering a surface that has animals on it. The sand (which will eventually turn into sandstone) covers the animals. Where the animal is tough, it will push into the overlying sand, causing a negative hyporelief impression (of the top surface of the animal) into the bottom surface of the sand. Where the animal is buried in the underlying sediment, such as holdfasts, the sand flows into the depression caused by the collapse of the holdfast during burial, causing a positive hyporelief impression.

The example above is of the lower surface, The Form D projects out from the surface and so is a positive hyporelief impression (the overlying sand has pushed down into the Form D stolon structure to form the impression. By contrast, the Parvancorina has pushed up into the overlying sand and so is preserved as a negative hyporelief impression.
OK, lets deal with the second line of evidence first - the possible stem extension emanating from a possible discoid fossil.

(A) Discoidal impression (negative relief) with possible stem-like extension. (B) Sketch showing the location of discoidal fossil and possible stem. Meert et al. 2010.
 Just to be clear about what is being discussed, here (at left) is a modern sea pen (photo credit) we can use as an example of the frondose forms around in the Ediacaran. The structure half-buried in the sediment is the disc-shaped holdfast. Underneath the holdfast is a thin tube - the peduncle - which acts like the foot of a clam and aids in digging the organism into the sediment. The stem and frond extend out into the water column and the whole thing is held up by water pressure. The organism can inflate by pumping water into the holdfast to allow it to act as an anchor, and also into the stem and frond to hold them rigid. Pumping water out deflates the organism. Modern sea pens will deflate on touching, and collapse down onto the sediment, as a method to avoid predation or damage.
Just to be clear about what is being discussed, here (at left) is a modern sea pen (photo credit) we can use as an example of the frondose forms around in the Ediacaran. The structure half-buried in the sediment is the disc-shaped holdfast. Underneath the holdfast is a thin tube - the peduncle - which acts like the foot of a clam and aids in digging the organism into the sediment. The stem and frond extend out into the water column and the whole thing is held up by water pressure. The organism can inflate by pumping water into the holdfast to allow it to act as an anchor, and also into the stem and frond to hold them rigid. Pumping water out deflates the organism. Modern sea pens will deflate on touching, and collapse down onto the sediment, as a method to avoid predation or damage.According to Meert et al., their find may represent a holdfast and part of the attached stem that has made an impression in the sediment, in negative relief. The authors do not say if the surface is the top or bottom (and they may not know if the sample was collected loose and not found in situ.
However, holdfast impressions are almost always found as positive hyporelief impressions, i.e. they stick out from the lower surface. They are not found as negative relief, i.e. as depressions, as the Kurgan Formation specimen is. To find out why that is, let's look at an example.

Three-dimensional positive hyporelief fossil of a holdfast and stem from the Ediacaran of South Australia. HF impression of holdfast, IS intermediary surface within the sandstone, LS lower surface of sandstone, P peduncle, S impression of stem, W wrinkles in top surface of holdfast. White arrow is the direction of current depositing the sand.Above is an actual Ediacaran holdfast and stem fossil. There's a lot going on here (compared with the Meert et al. specimen where no structure is present) so it'll take a bit of explaining.
Firstly observe that this is in positive hyporelief - it's on the bottom surface of the bed. As I said, pretty much all holdfast fossils with stems are positive hyporelief. This is because the organism is sitting in the underlying sediment and gets inundated and covered by a mantling sand deposit which will become the overlying sandstone bed. Since frondose forms will deflate on contact with the enveloping sandstone, the organism pumps out water and collapses. This means that the stem falls over, usually in the direction of sand transport. The stem often gets trapped within the sand and rarely comes to rest on the sea floor because the sand has started to deposit as the organism responds. This normally results in the stem coming to rest at an shallow angle to the sea floor/base of the sand body - that is, it sticks up at an angle into the sand rather than laying down flat. This can be seen in the Ediacaran example shown above. The stem impression (S) can be seen heading off at the ten o'clock direction but it is obviously heading off into the sandstone - the edge of the stem impression is lower in the sandstone than the impression of the holdfast, and, as this specimen is shown upside down, the stem in heading upwards into the overlying sandstone.
The wrinkling texture is the upper surface of the holdfast reacting to the stem being pushed over and collapsing and shows that the holdfast stayed in place during burial.
The organism is now covered by a layer of sand, with the stem trapped within the sand at a shallow angle. As the water has been pumped out, the holdfast will no longer stay inflated and so the holdfast upper surface sinks down onto the lower surface causing a shallow depression. The upper surface also takes on the shape of the lower surface where the hollow peduncle is, creating a ring shape (P). The overlying sand then flows into the shallow depression and takes on the shape of the combined lower and upper holdfast surface structures in a process called gravity casting. Also, as the stem is angled upwards, the sand with collect behind it and be trapped between the stem and the holdfast. We can see this is the example above. The sandstone covering the stem has been removed, leaving a lighter coloured sandstone. The stem starts out on top of the holdfast, but ends up on a lower, intermediate surface (IS) (lower from our prespective, higher when the specimen is rotated to the correct orientation).
What you end up with is a positive hyporelief impression of the combined upper and lower holdfast surface, plus the base of the stem. Furthermore, this is the only way this can be preserved. We tend not to see holdfast impressions on the tops of sandstones because the organisms appeared to favour softer, finer sediments such as silts and clays, and where active microbial mats occur.
With that in mind lets look at the Meert et al. specimen again.

This time I've annotated it to mark out the various levels in the specimen, with A the highest, B an intermediary level, and C the lowest (assuming we are looking at the top surface, or the reverse if we are looking at the lower surface).
First of all, this is a negative relief. As I explained above, holdfast and stem preservation is usually by gravity casting which produces positive relief. If we are looking at a bottom surface, maybe the organism remained intact and 'pumped up', forming a hollow in the overlying sand. But in that case the stem would also remain intact and we wouldn't see the impression of the stem in the resultant fossil. So if the stem is represented, then the organism had collapsed, in which case the holdfast would have collapsed and we would get a positive hyporelief as notmal.
The structure can't be a positive hyporelief that has eroded out the produce the depression because the original fossil is just a surface impression. The sandstone underneath the fossil is the same sandstone the rest of the rock and so there can be no preferential weathering to produce the hollow.
If we are looking at a top surface, then it is unlikely to be a holdfast and stem as we do not find them on top surfaces of sandstones. Even if it were, the stem appears to have been pushed into the sand body to a level equal to, or even greater than, the depth of the holdfast. Again an unlikely occurrence, as a slight impression on the underlying sand is the most you are likely to get.
Given that this is a negative relief and the lack of any distinguishing structure, it is most likely that the specimen is simply an erosional feature and not a fossil.
2)Nimbia and Aspidella
Specimens referable to Nimbia and Aspidella are reported from the c. 770 millio year old Kurgan Formation.
Our specimens were discovered in a brown-red shale within the largely siliclastic Kurgan Formation and are preserved in both hypo and epi-relief. They are circular to oval-shaped impressions that surround either a smooth interior or circular to oval indentations with a smooth interior. (Meert et al. 2010.)

(A) closeup of the Nimbia fossils in positive epirelief, AT marks the occurrence of a possible invaginate morph of Aspidella terranovica; (B) B&W photo of Nimbia impressions shown mostly in positive epirelief; (C) Nimbia fossils in negative relief with a possible invaginate morph of Aspidella terranovica in positive relief; (D) large c. 2 cm (long axis) c. 1.5 cm (short axis) negative hyporelief impression of Nimbia occlusa and an ‘inversion’ of the photo the fossil might appear in positive epirelief. White arrow points to a possible raised central nodule observed in Nimbia occlusa fossils elsewhere. Meert et al. 2010.Taking Aspidella first. Actually, attributing the form to Aspidella doesn't tell us much. This is because Aspidella should now be considered a form genus - that is a grouping of similar forms (i.e. discs) and not representative of a single organism, or related group of organisms. In other words, a lot of different things make disc impressions, e.g. algae (algal biscuits), microbial colonies, fungae and metazoans. Calling the specimen Aspidella simply says that it is a certain kind of disc, not that it is the Ediacaran form Aspidella terranovica. There are a lot of morphologies within the Aspidella group and they are not produced by the same organism, or even closely related organisms. This means that more than one organism is included under Aspidella. Even worse, no consistent distinction can be made between the various disk structures, making it difficult to accurately assign a particular disc to a particular organism. Where other evidence exists, i.e. stem impressions, ornamentation, etc, it is possible to identify individual organisms or groups of organisms. But simple discs remain probematic. Being a disc does not make it a holdfast!
It is entirely likely that Aspidella actually represents a group of diverse, not closely related, organisms - microbial, algal, fungal and metazoan - that produce discs and overlap each other to give Aspidella a large time range, but this time range is not for a single species or even genus.
This boils down to the fact that Aspidella with stems or other ornamentation, and associated with other Ediacaran forms, can be considered a metazoan fossil. Other Aspidella with no other ornamentation, and not associated with Ediacaran forms, could be anything.
The Nimbia find is in a similar situation. Do we have Nimbia? Yes. But here also, I think that the form Nimbia, like Aspidella, may hide a multitude of sins, or at the very least a number of different organisms. Except I think we have proof here that Nimbia is not an Ediacaran metazoan - or at least this Nimbia is not an Ediacaran metazoan.
The specimens of Nimbia from the Kurgan Formation are found as positive epirelief, negative epirelief, and negative hyporelief. This does not happen for Ediacaran fossils, as discussed above. Since these Nimbia show a number of modes of preservation, and they all show the same structure regardless of mode of preservation, it is apparent that the form that produced it is rigid. It has to be if it produced the same shape whether on the top surface creating a positive or negative imprints, or on the bottom surface. The fossil is the same independent of preservational style, and so whatever is causing the fossil must therefore be rigid like a coin. That rigidity, and the various preservational styles, means that it cannot be a holdfast.
It has been suggested that Nimbia represents microbial colonies. Hard, well agglutinated disk-shaped colonies would be tough enough to cause the impressions, and some have been known to develop a central nodule similar to that found in some Nimbia. This would explain the similar shape through different modes of preservation, and the large time range of the form.
Finally, Ediacaran fossils are normally found as assemblages, with a number of different forms present. The Kurgan Formation specimens are of one main type, Nimbia, and a few Aspidella. Too restricted to be considered an assemblage (a random collection would be expected to uncover more than two forms if an assemblage was present). Even if this find represented an early stage in the evolution of metazoa, it would be expected that more than two forms would be present.
So, discoial fossil with stem? - no. Aspidella? - yes, possibly, but that doesn't mean it's an Ediacaran or even metazoan. Nimbia? - yes, but probably not metazoan.
In properly posing questions about their finds instead of making pronouncements, and suggesting a number of possible interpretations for their finds, Meert at al. have highlighted the important implication of their finds.
It is possible that our discovery of Nimbia occlusa and Aspidella terranovica(?) in sedimentary rocks during the early Cryogenian (N766 Ma) lends support the alternative hypotheses regarding these fossils and remove them from consideration as true metazoa(Meert et al. 2010.)I think they are right, they have provided evidence that the forms are not metazoan - or at least not "Ediacaran" (as in found amongst Ediacaran assemblages) - fossils, and that the simple disc-shaped forms found in the Proterozoic comprise a form group with contributions from a number of metazoan and non-metazoan sources.
Meert, J., Gibsher, A., Levashova, N., Grice, W., Kamenov, G., & Ryabinin, A. (2010). Glaciation and ~770Ma Ediacara (?) Fossils from the Lesser Karatau Microcontinent, Kazakhstan Gondwana Research DOI: 10.1016/j.gr.2010.11.008
Further reading on Ediacaran discs
Gehling, J.G., Narbonne, G.M., and Anderson, M.M (2000) The first named ediacaran body fossil Aspidella terranovica. Palaeontology, 43: 427-456. DOI: 10.1111/j.0031-0239.2000.00134.x
MacGabhann, B.A. (2007) Discoidal fossils of the Ediacaran biota: a review of current understanding. Geological Society, London, Special Publication, 286: 297-313. DOI: 10.1144/SP286.21
Wednesday, December 29, 2010
The Palaeontological Twelve Days of Christmas - Day 12
Following on from the Twelve Geology Days of Christmas, this year we have a palaeontological version.
On the twelfth day of Christmas my true love sent to me, twelve insects in Australian amber entombing,
eleven ediacaran organisms evolving,
ten tiny T. rex tyrranizing,
nine Nigersaurus nibbling,
eight evolutionary digits encroaching,
seven seismosaurus stomping,
six snakes aslithering,
fiiiiive fossilllll fishhhhhhh,
four fossil footprints following,
three Tribrachidium tripartiting
two trilobites tasting,
and a Cambrian Lagerstätte.
Subscribe to:
Posts (Atom)