On the twelfth day of Christmas my true love sent to me, twelve insects in Australian amber entombing,
eleven ediacaran organisms evolving,
ten tiny T. rex tyrranizing,
nine Nigersaurus nibbling,
eight evolutionary digits encroaching,
seven seismosaurus stomping,
six snakes aslithering,
fiiiiive fossilllll fishhhhhhh,
four fossil footprints following,
three Tribrachidium tripartiting
two trilobites tasting,
and a Cambrian Lagerstätte.
Wednesday, December 29, 2010
The Palaeontological Twelve Days of Christmas - Day 12
Following on from the Twelve Geology Days of Christmas, this year we have a palaeontological version.
Tuesday, December 28, 2010
The Palaeontological Twelve Days of Christmas - Day 11
Following on from the Twelve Geology Days of Christmas, this year we have a palaeontological version.
On the eleventh day of Christmas my true love sent to me, eleven ediacaran organisms evolving,
ten tiny T. rex tyrranizing,
nine Nigersaurus nibbling,
eight evolutionary digits encroaching,
seven seismosaurus stomping,
six snakes aslithering,
fiiiiive fossilllll fishhhhhhh,
four fossil footprints following,
three Tribrachidium tripartiting
two trilobites tasting,
and a Cambrian Lagerstätte.
Monday, December 27, 2010
The Palaeontological Twelve days of Christmas - Day 10
Following on from the Twelve Geology Days of Christmas, this year we have a palaeontological version.
On the tenth day of Christmas my true love sent to me, ten tiny T. rex tyrranizing,
nine Nigersaurus nibbling,
eight evolutionary digits encroaching,
seven seismosaurus stomping,
six snakes aslithering,
fiiiiive fossilllll fishhhhhhh,
four fossil footprints following,
three Tribrachidium tripartiting
two trilobites tasting,
and a Cambrian Lagerstätte.
Sunday, December 26, 2010
The Palaeontological Twelve Days of Christmas - Day 9
Following on from the Twelve Geology Days of Christmas, this year we have a palaeontological version.
On the ninth day of Christmas my true love sent to me, nine Nigersaurus nibbling,
eight evolutionary digits encroaching,
seven seismosaurus stomping,
six snakes aslithering,
fiiiiive fossilllll fishhhhhhh,
four fossil footprints following,
three Tribrachidium tripartiting
two trilobites tasting,
and a Cambrian Lagerstätte.
Saturday, December 25, 2010
The Palaeontological Twelve Days of Christmas - Day 8
Following on from the Twelve Geology Days of Christmas, this year we have a palaeontological version.
On the eighth day of Christmas my true love sent to me, eight evolutionary digits encroaching,
seven seismosaurus stomping,
six snakes aslithering,
fiiiiive fossilllll fishhhhhhh,
four fossil footprints following,
three Tribrachidium tripartiting
two trilobites tasting,
and a Cambrian Lagerstätte.
Friday, December 24, 2010
Why Dinosaurs Hate Christmas
Back by popular demand, and new and improved (well it has a new diagram . . .)
Did you know that most dinosaurs hate Christmas? It’s true, they do. And it’s not because they couldn’t get a handle on the present wrapping (or unwrapping for that matter) either. No, there is a very good reason why Dinosaurs hate Christmas.
 However, before explaining why dinosaurs hate Christmas, lets deal with some startling new information. Everyone is familiar with the standard explanation for the extinction of the dinosaurs. I've included a common representation of the event.
However, before explaining why dinosaurs hate Christmas, lets deal with some startling new information. Everyone is familiar with the standard explanation for the extinction of the dinosaurs. I've included a common representation of the event.
However, startling 'evidence' has been presented which suggests another reason for what happened. The evidence is still officially hidden by the authorities, but one startling image has been smuggled out and is shown here for the first time.
It is claimed that it was one of Santa's early experiments on propulsion systems that went wrong and had to be ejected. Shocking as this image is, there are some who claim that it is a forgery and just another shot by those at war with Christmas.
However, this is not the reason that dinosaurs hate Christmas. To understand that we need to know what dinosaurs are.
In his classic 1842 publication on dinosaurs, Richard Owen named and defined the Dinosauria as:
a group of exceedingly large, pachydermous reptiles from the Second Age . . . includes Megalosaurus, Iguanodon and Hylaeosaurus.
In 1997, Tom Holtz provided a different definition:
the last common ancestor of Megalosaurus andIguanodon and all its descendants. Using the same methodology, we can define dinosaurs as including the last common ancestor of the Herrerasauidae and the Hadrosauridae, and all their decendents.

Anyhow, the thing about this definition is that, nestled between the the Herrerasaurs and the Hadrosaurs, are the Dromaeosaurs, and directly related to the Dromaeosaurs, and so one of the descendants mentioned above, is a little group called Aves!
So with the mass slaughter ofbirds dinosaurs every Christmas, wouldn't you hate Christmas?
If you insist in participating in this slaughter, at least make sure you cook your dinosaur correctly:
1. If your dinosaur is frozen, fully thaw it.
2. Don’t stuff the dinosaur. By the time the stuffing reaches a safe temperature, the meat is overcooked.
3. Cover the dinosaur breast with ice while the rest of the dinosaur warms to room temperature. Don’t leave the dinosaur out for more than 3 hours. At this point, the breast will be about 4.5oC (40oF), while the rest of the meat will be at 15.5oC (60oF).
4. Put the dinosaur in the oven and cook according to your favorite recipe.
5. With a meat thermometer, check temperature. Take out of the oven when legs reach 82oC(180oF) and breast hits between 68-71oC (155-160oF).
Ho Ho Bleedin' Ho.
Did you know that most dinosaurs hate Christmas? It’s true, they do. And it’s not because they couldn’t get a handle on the present wrapping (or unwrapping for that matter) either. No, there is a very good reason why Dinosaurs hate Christmas.
 However, before explaining why dinosaurs hate Christmas, lets deal with some startling new information. Everyone is familiar with the standard explanation for the extinction of the dinosaurs. I've included a common representation of the event.
However, before explaining why dinosaurs hate Christmas, lets deal with some startling new information. Everyone is familiar with the standard explanation for the extinction of the dinosaurs. I've included a common representation of the event.However, startling 'evidence' has been presented which suggests another reason for what happened. The evidence is still officially hidden by the authorities, but one startling image has been smuggled out and is shown here for the first time.
It is claimed that it was one of Santa's early experiments on propulsion systems that went wrong and had to be ejected. Shocking as this image is, there are some who claim that it is a forgery and just another shot by those at war with Christmas.
However, this is not the reason that dinosaurs hate Christmas. To understand that we need to know what dinosaurs are.
In his classic 1842 publication on dinosaurs, Richard Owen named and defined the Dinosauria as:
a group of exceedingly large, pachydermous reptiles from the Second Age . . . includes Megalosaurus, Iguanodon and Hylaeosaurus.
In 1997, Tom Holtz provided a different definition:
the last common ancestor of Megalosaurus andIguanodon and all its descendants. Using the same methodology, we can define dinosaurs as including the last common ancestor of the Herrerasauidae and the Hadrosauridae, and all their decendents.

Anyhow, the thing about this definition is that, nestled between the the Herrerasaurs and the Hadrosaurs, are the Dromaeosaurs, and directly related to the Dromaeosaurs, and so one of the descendants mentioned above, is a little group called Aves!
So with the mass slaughter of
If you insist in participating in this slaughter, at least make sure you cook your dinosaur correctly:
1. If your dinosaur is frozen, fully thaw it.
2. Don’t stuff the dinosaur. By the time the stuffing reaches a safe temperature, the meat is overcooked.
3. Cover the dinosaur breast with ice while the rest of the dinosaur warms to room temperature. Don’t leave the dinosaur out for more than 3 hours. At this point, the breast will be about 4.5oC (40oF), while the rest of the meat will be at 15.5oC (60oF).
4. Put the dinosaur in the oven and cook according to your favorite recipe.
5. With a meat thermometer, check temperature. Take out of the oven when legs reach 82oC(180oF) and breast hits between 68-71oC (155-160oF).
Ho Ho Bleedin' Ho.
The Palaeontological Twelve Days of Christmas - Day 7
Following on from the Twelve Geology Days of Christmas, this year we have a palaeontological version.
On the seventh day of Christmas my true love sent to me, seven seismosaurus stomping,
six snakes aslithering,
fiiiiive fossilllll fishhhhhhh,
four fossil footprints following,
three Tribrachidium tripartiting
two trilobites tasting,
and a Cambrian Lagerstätte.
Thursday, December 23, 2010
The Palaeontological Twelve Days of Christmas - Day 6
Following on from the Twelve Geology Days of Christmas, this year we have a palaeontological version.
On the sixth day of Christmas my true love sent to me, six snakes aslithering,
fiiiiive fossilllll fishhhhhhh,
four fossil footprints following,
three Tribrachidium tripartiting
two trilobites tasting,
and a Cambrian Lagerstätte.
Wednesday, December 22, 2010
The Palaeontological Twelve Days of Christmas - Day 5
Following on from the Twelve Geology Days of Christmas, this year we have a palaeontological version.
On the fifth day of Christmas my true love sent to me,
fiiiiive fossilllll fishhhhhhh,
four fossil footprints following,
three Tribrachidium tripartiting
two trilobites tasting,
and a Cambrian Lagerstätte.
Tuesday, December 21, 2010
The Palaeontological Twelve Days of Christmas - Day 4
Following on from the Twelve Geology Days of Christmas, this year we have a palaeontological version.
On the fourth day of Christmas my true love sent to me, four fossil footprints following,
three Tribrachidium tripartiting
two trilobites tasting,
and a Cambrian Lagerstätte.
Monday, December 20, 2010
The Palaeontological Twelve Days of Christmas - Day 3
Following on from the Twelve Geology Days of Christmas, this year we have a palaeontological version.
On the third day of Christmas my true love sent to me, three Tribrachidium tripartiting,
two trilobites tasting,
and a Cambrian Lagerstätte.
Sunday, December 19, 2010
The Palaeontological Twelve Days of Christmas - Day 2
On the second day of Christmas my true love sent to me, two trilobites, and a Cambrian Lagerstätte.
Saturday, December 18, 2010
The Palaeontological Twelve Days of Christmas - Day 1
Following on from the Twelve Geology Days of Christmas, this year we have a palaeontological version.
On the first day of Christmas my true love sent to me, a Cambrian Lagerstätte.
Monday, November 22, 2010
Palaeoporn 20

Frondose (frond-bearing) forms are the most complex and interesting of the Ediacaran fossils, but rare. This is because most of the organism lives above the sea floor, to which they are anchored by a round holdfast, and so are not preserved. A number of frondose forms have been found, but Pambikalbae is different.
 Pambikalbae hasenohrae (a) complete block showing several specimens (upper right and lower left). Slab 45 cms. (b) same slab but tilted to show fossil is interlayers with the sandstone matrix (cracks on left side of slab). (c) part of the right hand frond in (a) showing a complex arrangement of chambers.
Pambikalbae hasenohrae (a) complete block showing several specimens (upper right and lower left). Slab 45 cms. (b) same slab but tilted to show fossil is interlayers with the sandstone matrix (cracks on left side of slab). (c) part of the right hand frond in (a) showing a complex arrangement of chambers.Pambikalbae was made up of numerous chambered vanes, making up a frond, supported by a tapering central stem and an anchoring stalk. Several series of chambers occured on the vanes, joined together at zigzag sutures, and were commonly filled with sediment on burial. However, the complexity is probably an artifact of complex composite moulding of various nearby chambers, one on the other. It appears that the chambers curve away from the central stem out to the free, or outer, margin. The chambers also appear to be set on the vane in a regular patter so as to limit overlap.
These chambers are big. Bigger than anything else so far found. Here's a reconstruction.

Pambikalbae is clearly not a pennatulacean or cnidarian 'sea pen' like Charnodiscus. However, it does contain characters that suggest an evolutionary grade of organisation comparable to known cnidarians. The configuration and size of the chambers seems ideal to house symbiotic algae or bacteria . . . however, here's some wild speculation.
There is a group of cnidarians which do share similarities with Pambikalbae, and that's the physophorid Siphonophorida. Here's one below (photo credit)

Interesting isn't it. The chambers are especially similar. However, there is one thing wrong. The physophorid is upside down. The chambers actually point downwards. Why? Because physophorids float, they are not anchored to the sea floor like Pambikalbae.

But it's not as problematic getting from Pambikalbae to a physophorid, as it would appear.
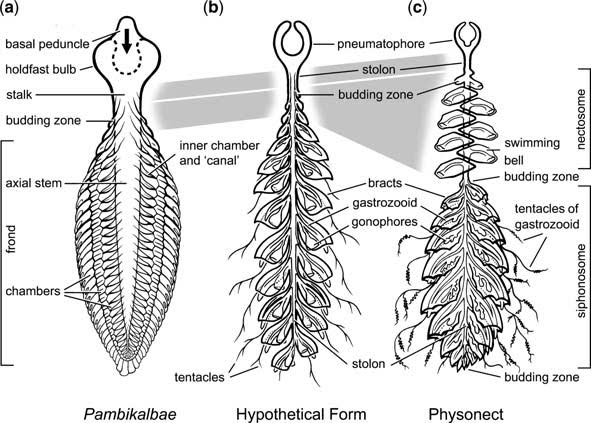 (a) inverted Pambikalbae. (b) a hypothtical ancestral 'calycophore' siphonophore. (c) generalised modern physonect.
(a) inverted Pambikalbae. (b) a hypothtical ancestral 'calycophore' siphonophore. (c) generalised modern physonect.Just change a water-filled bulb as an holdfast, for an air filled bulb for a float. Simple folding inwards of the holdfast could produce the physonect and ancestral calycophore pneumatophore. A futher point of comparison is that the vane of Pambikalbae has three serial rows of chambers, as in calycophore and physonect siphonophores.
So a frondose form with wanderlust as ancestral to the modern siphonophore cnidarians? Maybe. But is could also be a derived hyrozoan, or a sister group to the early Chondrophorina. Whatever it is, it ain't no pennatulacean!
Jenkins, R.J.F. and Nedin, C. (2007) The provenance and palaeobiology of a new multi-vane, chambered frodose organism from the ediacaran (later Neoproterozoic) of South Australia. In P. Vickers-Rich, P. and Komarower, P. (eds) The Rise and Fall of the Ediacaran Biota. Geological Society Special Publication 286, 195-222. doi: 10.1144/SP286.15
Saturday, November 6, 2010
The Royal Society Archive and Optick Glaffes
The Royal Society is 350 years old in 2010. To mark the event, the Royal Society archives have been made freely available online.
ALL of them.
From 1665.
350 years worth.
The archives contain more than 66,000 articles, including the first ever article published in the world's first science journal Philosophical Transactions.

Here's a nerdy quiz question for you to ask, "What was the subject of the first paper in the first scientific journal?" Answer - Optick Glaffes.
The archive will remain free to access until 30 November 2010. So go in now and download some original articles from people such as Mr Issac Newton (a promising young mathematiks Profeffor)

and a Mr Charles Darwin (recently back from an around-the-world cruise).

ALL of them.
From 1665.
350 years worth.
The archives contain more than 66,000 articles, including the first ever article published in the world's first science journal Philosophical Transactions.

Here's a nerdy quiz question for you to ask, "What was the subject of the first paper in the first scientific journal?" Answer - Optick Glaffes.
The archive will remain free to access until 30 November 2010. So go in now and download some original articles from people such as Mr Issac Newton (a promising young mathematiks Profeffor)

and a Mr Charles Darwin (recently back from an around-the-world cruise).

Friday, November 5, 2010
Lower Cambrian Sea Anemones from China
 Holotype and paratypes of Eolympia pediculata from Han et al. 2010
Holotype and paratypes of Eolympia pediculata from Han et al. 2010Yet more exquisitely preserved fossils from the phosphorite deposits in the lowest Cambrian sediments of the Kuanchuanpu Formation, Shaanxi, China. And by "lowest" they really mean lowest! The deposits are only a couple of million years younger that the Cambrian-PreCambrian boundary, which is currently taken as 542 million years ago.
The new fossils have been identified as a possible stem member of the Cnidarian Hexacorallia, suggesting that the diversification of the Cnidaria either occurred very rapidly after the start of the Cambrian, or, more likely (as far as I am concerned), in the Ediacaran.
I don't have much comment to make. The interpretation appears reasonable. The paper is freely available at PLoS (thank you PLoS). I would have liked some larger specimens, but the size is an artifact of the preservation.
There's a nice comparison with some extant polyps from an extant species.
 Young polyps from a modern species, from Han et al. 2010
Young polyps from a modern species, from Han et al. 2010The similarity in form and size is striking. Morphological similarity isn't everything, but it's something!
The authors end with:
The cnidarian diversification might have occurred rather quickly during the early half of the Cambrian or it may be deeply rooted into the Neoproterozoic.I prefer the latter option, which is a nice intro to Palaeoporn 20!
Han J, Kubota S, Uchida H-o, Stanley GD Jr, Yao X, et al. (2010) Tiny Sea Anemone from the Lower Cambrian of China. PLoS ONE 5(10): e13276. doi:10.1371/journal.pone.0013276
Monday, October 11, 2010
Palaeoporn 19
Another holy grail

Ignore the cute baby buffalo. See that unprepossessing roadside quarry with the people sitting on a pile of rubble (click to enlarge)? Well that rubble is the type section for the Chengjiang fauna.
Yeah! Not so unprepossessing now, is it?!
The Chengjiang fauna occurs in the Maotianshan Shale, a member of the Lower Cambrian Chiungchussu formation. They are found about 5 km northwest of the Fuxian Lake, and 6 km northeast of Chengjiang, Yunnan, China.
The age of the fauna is between 525-520 million years old, significantly older than the Burgess Shale (at 505 million years ago), and the Emu Bay Shale (approx 515 million years old).
The Chengjiang fauna rivals that of the Burgess Shale in preservation and diversity. Although not in difficulty to get to. The Burgess Shale is a 3 hour hike. Here, you can drive right to the outcrop!

In this shot (click to enlarge) you can see the quarry from photo one in the middle distance. There is a bus parked next to it with some people in the road. The hill to the right is Maotian Mount (hence Maotianshan Shale)

In this photo (click to enlarge), I am right at the back in short sleeves and a broad-brimmed brown hat (I'm in the same position in the first photo). I'm there partly because it's in the shade, and partly because there were some nice fossils there (all went to Nanjing University for study).
The depositional environment was delta front prograding eastwards into an open sea. Most of the fossil layers appear to be episodic events onto the marine muds in front of the delta, possible storm induced deposition of clays and fine sand. There's not much evidence of transport, so most organisms were locals and were buried by a series of turbidity flows.

This is a smaller quarry close by (click to enlarge). The dark colour is the fresh colour of the Maotianshan Shale. It is black when fresh, but rapidly oxidises to a tan colour on exposure to air. This is near the top of the Maotianshan Shale. The pick axe handle in the middle of the shot is marking the topmost Maotianshan Shales. The very top of the handle is resting against the first influx of sands, which coarsen upward until they are topped by a large lenticular sandstone at the top of the sequence above my head. This represents the prograding delta as it moves out over the muds of the Maotianshan Shale - just like the sands of the Mississippi Delta are prograding out into the Gulf of Mexico.
Monday, October 4, 2010
Evolution Among the Trilobites - Part 2

In Part 1 we looked at the growth patterns of the Early Cambrian trilobite Estaingia bilobata, using certain measurements from the head, or cranidium. In this part we'll compare and contrast those growth patterns with the Early Cambrian trilobite Xystridura templetonensis. You can click on any of the images and graphs to get a larger version.
The change through time in the appearance or rate of development of ancestral characters is known as Heterochrony, which comprises two basic phenomena (pay attention, there'll be a quiz at the end!):
Paedomorphosis - the retention of ancestral juvenile characteristics in a descendant adult. Paedomorphic forms usually pass through fewer morphological stages during growth than their ancestors (in other words less time as a juvenile you stop being a juvenile early, before you've shed all the baby characters). There are three types of peadomorphosis
Deceleration - the rate of morphological development is reduced during the juvenile phase (slower development, but the same amount of time is spent as a juvenile, so the same size as the ancestor, but you have not completed the juvenile development so have a baby-faced adult)Peramorphosis - the appearance of ancestral adult features in the descendant juvenile stage (beards on a baby!). Here to there are three processes:
Hypomorphosis - the onset of maturity occurs at an earlier stage of development (same development but less time as a juvenile, so a smaller adult then the ancestor - still baby-faced though).
Post-displacement - a change in the timing of the onset of certain features, with one or more structures starting to develop at a later stage (the same amount of time is spent as a juvenile, so the same size as the ancestor, but the structures are smaller in the adult)
Acceleration - increasing the rate of morphological development (if maturity is also accelerated then adult will be smaller then the ancestral adult, if the onset of maturity is not affected, the the adult will be the same size as the ancestral adult).
Hypermorphosis - delayed maturity so the juvenile stage is extended (spend longer as a juvenile, so the adult is bigger)
Pre-displacement development of structures occurs at an earlier stage of development (same time as a juvenile, but structures more developed and bigger).
.jpeg) Before we look at a comparison between Estaingia and Xystridura, lets refresh what it is we are actually measuring (see Estaingiacranidium (or head) at right). The relevant measurements that we are interested in are; Cranidial Width (CW), which is literally the distance between the eyes; the Pre-glabella Field (PGF), which is the area in front of the large bulbous glabella (incidentally the pre-glabella field only runs to the shallow trench towards the front of the cranidium. In front of the trench is the doublure. That is folded underneath the cranidium in life, along the trench, and pops up during moulting); Glabella Length (GL), adding PGF and GL gives us a value for the length of the cranidium; Pre-Orbital Glabella (POG), which is the bit of the glabella that lies in front of a line drawn between the front tip of the eyes; and the distance between the end of the Axial (or Occipital) Furrow (the trench behind the glabella) and the back tip of the Eye Lobe (AF-EL). (Note that the figure is missing the free cheeks. This is because they are usually lost or repositioned during moulting. Trying to measure missing of displaced portion of the head would not allow accurate measurements. The measurements here, therefore, are all done on cranidia (without free cheeks).
Before we look at a comparison between Estaingia and Xystridura, lets refresh what it is we are actually measuring (see Estaingiacranidium (or head) at right). The relevant measurements that we are interested in are; Cranidial Width (CW), which is literally the distance between the eyes; the Pre-glabella Field (PGF), which is the area in front of the large bulbous glabella (incidentally the pre-glabella field only runs to the shallow trench towards the front of the cranidium. In front of the trench is the doublure. That is folded underneath the cranidium in life, along the trench, and pops up during moulting); Glabella Length (GL), adding PGF and GL gives us a value for the length of the cranidium; Pre-Orbital Glabella (POG), which is the bit of the glabella that lies in front of a line drawn between the front tip of the eyes; and the distance between the end of the Axial (or Occipital) Furrow (the trench behind the glabella) and the back tip of the Eye Lobe (AF-EL). (Note that the figure is missing the free cheeks. This is because they are usually lost or repositioned during moulting. Trying to measure missing of displaced portion of the head would not allow accurate measurements. The measurements here, therefore, are all done on cranidia (without free cheeks).The story is that Xystridura (Middle Cambrian) has evolved from Estaingia as represented by forms from the Emu Bay Shale (Lower Cambrian) through changes in the rate of development of certain characters, or hererochrony.
Now, to look at changes over time we will be comparing ratios, not individual characters. Individual characters will usually grow over time, but we are interested in how that character changes with growth. If the character increased with growth at a greater rate than other characters, then the ratio increases. If the character increases with growth at a lesser rate than other characters, then the ratio decreases. If the character increases at the same rate as other characters then there is no change in the ratio.
The first character we will look at is the distance between the back of the eye and the occipital furrow (AF-EL).
.jpeg) Measurements of meraspid Estaingia and Xystridura.
Measurements of meraspid Estaingia and Xystridura. Xystridura measurements adapted from McNamara (1981).
Here, the ratio AF-EL/CL compared with cranidial length is plotted with growth (cranidial length is taken as a proxy for growth) for meraspid (juvenile) Estaingia (yellow) and Xystridura (black). Here the smallest (youngest) meraspids have the largest ratio, and as they grow that ratio decreases. This means that, with growth, the feature is increasing at a slower rate than the overall growth rate. We can also see that Estaingia and Xystridura plot together. This means that the juvenile growth pattern is the same for both. The big difference is that the Xystridura meraspids continue beyond the transition from Estaingia meraspid to holaspid (adult) (yellow dotted line). The Xystridura meraspid to holaspid transition (black dotted line) occurs later than Estaingia and so Xystridura meraspids grow to a larger size before the holaspid stage than do Estaingia meraspids. So the onset of maturity is delayed and the juvenile growth phase has been extended.
The largest Xystridura meraspid appear to have a smaller AF-EL/CL ratio than Estaingia meraspids, but the spread of data is large.
Obviously, the character itself - the distance between the back of the eye ridge and the occipital furrow - increases throughout growth in absolute terms, but that increase is at a slower rate that the overall growth of other characters, so the ratio decreases.
No lets look at what happens during holaspid growth.
.jpeg) Measurements of Estaingia and Xystridura.
Measurements of Estaingia and Xystridura. Xystridura measurements adapted from McNamara (1981).
The first thing to note is that the meraspid growth pattern of reducing AF-EL/CL ratio is halted at the meraspid-holaspid transition. In the holaspid growth phase the AF-EL/CL ratio does not change with growth, which means that it is now increasing at the same rate as the length of the head. Also the spread of data at the meraspid-holaspid transition for Xystridura meraspid settles down in the holaspids suggesting that the timing of the transition has a bit of slop in it.
The second thing to note is that Estaingia and Xystridura have the same growth pattern - exactly the same growth pattern. Having a similar growth pattern could be chance - maybe this is a common growth pattern for trilobites. But having exactly the same ratio values is unlikely to occur by chance. This is strong evidence of an evolutionary relationship. Xystridura has inherited this particular ratio pattern and values from Estaingia.
Note also the "NSW Estaingia". This is Estaingia bilobata from Cymbric Vale in New South Wales. It is slightly younger than the Estaingia bilobata from the Emu Bay Shale. There are few measurements, but this form plots on the same trend and ratio values as the others - albeit a larger form than the Estaingia bilobata from the Emu Bay Shale.
Lets look at another character, this time the ratio of width compared with length of the head CW/CL
.jpeg) Measurements of meraspid Estaingia and Xystridura.
Measurements of meraspid Estaingia and Xystridura. Xystridura measurements adapted from McNamara (1981).
Here again are the meraspid measurements for Estaingia and Xystridura. Again the character ratio decreases with growth. This means that as the head grows, the width is growing at a slower rate than the length. Also, the Xystridura meraspid growth phase is extended. This time however, there does seem to be some difference between the Estaingia and Xystridura meraspid growth phases. Unlike the last example, where the two growth patterns seemed to be in step. here the Xystridura meraspids appear to change the growth pattern. The larger Xystridura meraspids appear to stop the trend of decreasing CW/CL ratio and, by the time they reach the meraspid-holaspid transition, they appear to be slightly increasing the CW/CL ratio.
Things should become clearer when we look at the complete growth patterns.
.jpeg) Measurements of Estaingia and Xystridura.
Measurements of Estaingia and Xystridura. Xystridura measurements adapted from McNamara (1981).
Here the growth patterns are markedly different. It should be pointed out that holaspid growth changes usually occur at slower rates than for meraspid forms, so that holaspid trends are less marked than meraspid trends. The Estaingia growth pattern shows that the reduction in the CW/CL ratio occurring in the meraspid growth phase is slowed at the meraspid-holaspid boundary. In the holsapid growth phase, the growth pattern shows that cranidial width continues to grow at a slower rate than cranidial length, but the difference between the growth rates is reduced somewhat.
However, for Xystridura, the apparent change in growth rate that occurs in the latest meraspids is carried into the holaspid growth phase, where the CW/CL ratio not only equalises (that is the width is growing at the same rate as length) but the width growth rate actually appears to be slightly exceeding the length growth rate.
Critically this change in Xystridura growth pattern occurs in the meraspid phase, but at the point where Estaingia would be expected to transition into the holaspid growth pattern. It isn't very clear in the graph (so click on it to enlarge) but the new growth pattern of the larger Xystridura meraspid after the Estaingia meraspid-holaspid transition (yellow dotted line) shows that the width is actually growing faster than the length (CW/CL ratio is increasing). At the Xystridura meraspid-holaspid transition (black dotted line) this growth rate actually slows down so that the width growth rate only slightly exceeds the length growth rate.
The NSW Estaingia actually plots between the Estaingia and Xystridura, suggesting that this change was underway by the time of Cymbric Vale sediments deposition.
The one thing that controls the cranidial width is the large bulbous thing in the centre of the head - the glabella. What these measurements are telling us is that during Estaingia meraspid growth, the glabella increases in width at a slower rate the the increase in the length of the head. During holaspid growth, the rate of growth of glabella width actually increases, but still not to the rate of increase of the length of the head - the glabella has increased the pace of its growth, but not enough to match the rate of growth in length the head is achieving - resulting in a slowing of the trend that is reducing the ratio of width to length, but not enough to stop the trend. In Xystridura, the glabella width actually starts to grow at a faster rate than the length - it is increasing in width ata faster rate than the head is growing longer - and so the ratio starts to increase.
If the glabella is changing size in one direction, maybe it's changing in another. Lets see.
The length of the head is composed of two measurements, the glabella. and the field in front of the glabella - the Preglabella Field (PGF) Any change in the glabella will affect the PGF. If the glabella increases in size at a faster rate than the length of the cranidium, then the PGF will reduce, if the glabella increases in size at a slower rate than the length of the cranidium, then the PGF will increase. If the glabella increase in size at the same rate as the length of the cranidium, then the PGF will remain the same. Measuring the PGF then, provides a proxy for glabella length growth.
.jpeg) Measurements of meraspid Estaingia and Xystridura.
Measurements of meraspid Estaingia and Xystridura. Xystridura measurements adapted from McNamara (1981).
For Estaingia meraspid, the PGF is increasing with increasing meraspid size. This means that the glabella is growing at a slower rate then the length of the head resulting in an expanded PGF. By the Estaingia meraspid-holaspid transition, this pattern has stopped.
The Xystridura meraspid growth pattern follows the Estaingia meraspid pattern in showing an increase in the size of the PGF up to a certain point. Then, just before the Estaingia meraspid-holaspid transition this trend reverses, and the PGF reduced in size. This means that the glabella is now increasing in size at a greater rate than the length of the cranidium, resulting in a reduction in the PGF.
What does that mean for the holaspids?
.jpeg) Measurements of Estaingia and Xystridura.
Measurements of Estaingia and Xystridura. Xystridura measurements adapted from McNamara (1981).
Lets take Estaingia first. After the meraspid-holaspid transition, the PGF ratio decreases slowly. This indicated that the glabella is growing at a faster rate than the cranidial length, so the PGF is getting smaller, but only slowly.
After the Xystridura meraspid-holaspid transition, Xystridura shows the same pattern of slowly reducing PGF and so slowly increasing glabella. But, as the Xystridura meraspid phase lasted longer, the trend to reducing PGF (increasing glabella) starts in the meraspid period and is accentuated. So much so that, by the time of the Xystridura meraspid-holaspid transition, the PGF is so small that even at the slowly reducing rate of the holaspid form, the PGF reached zero shortly after the Xystridura meraspid-holaspid transition. This means that the the glabella has been growing at such a faster rate than the cranidium, that it is so close to the front of the cranidium by the time of the meraspid-holaspid transition, that it quickly reaches the front of the cranidium early in the holaspid phase. PGF = 0.
Interestingly, the NSW Estaingia plot with the other Estaingia. This indicated that this change had not yet started by Cymbric Vale time.
 All it took to produce a descendant form that looks significantly different from the ancestral form is the simple process of slightly delaying the onset of the holaspid phase, resulting in some of the slower holaspid growth pattern being incorporated into the faster meraspid pattern. In other words the large bulbous glabella of Xystridura is caused by a portion of the holaspid growth pattern being incorporated into the meraspid growth phase. "But wait a minute", you say (or you would say if you were paying attention) "the first character didn't change at all, and that involves the glabella - it measures the distance between the back of the eye ridge and the side of the back end of the glabella". That's true, but the growth of the glabella is focused towards the front of it. It is the front portion that has grown, not the back portion. So the back remains unaffected by the change.
All it took to produce a descendant form that looks significantly different from the ancestral form is the simple process of slightly delaying the onset of the holaspid phase, resulting in some of the slower holaspid growth pattern being incorporated into the faster meraspid pattern. In other words the large bulbous glabella of Xystridura is caused by a portion of the holaspid growth pattern being incorporated into the meraspid growth phase. "But wait a minute", you say (or you would say if you were paying attention) "the first character didn't change at all, and that involves the glabella - it measures the distance between the back of the eye ridge and the side of the back end of the glabella". That's true, but the growth of the glabella is focused towards the front of it. It is the front portion that has grown, not the back portion. So the back remains unaffected by the change.Why is the glabella important? Well, that's where trilobites keep the stomach. So big glabella, big stomach.
So, what type of heterochrony is this? Go back to the list at the start and work it out.
I'll give you a clue. We have delayed maturity so the juvenile stage is extended. As a result, the the adult is bigger.
We are dealing here with Peramorphosis, and more specifically Hypermorphosis. One or more populations of Estaingia has evolved, by Hypermorphosis into Xystridura. An event that occurred around the Lower-Middle Cambrian boundary, during a time of eustatic sea level fall that would have reduced shallow-water living space.
The take-home message isn't that trilobites are cool (they are), but that this brings out a very important point about evolution, and a good refutation of the old creationist canard, "if evolution is true where are the half-way transitionals? The half reptile-half bird?"
What these results show, is that evolution doesn't happen to all features at the same time, or at the same rate, producing a neat half-and-half transitional form. Some features change relatively rapidly (the expansion of the frontal glabella), some features change relatively slowly (head width to length ratio), and some don't change at all (the distance from the back of the eye to axial furrow distance as a ratio of head length). So there isn't a transitional which has all features exactly half way between the ancestral and descendant forms. What we find are transitionals with a mix of features depending on the rates at which those features are changing. We should not expect to find exact half-and-half transitionals. Evolution doesn't work that way.
de Beer called it "Mosaic Evolution". That isn't an excuse for the lack of half-and-half transitionals, it's a description of how evolution operates.
McNamara, K.J. (1981) Paedomorphism in Middle Cambrian xystridurine trilobites from northern Australia. Alcheringa, 5: 209-224. DOI: 10.1080/03115518108567002
Sunday, September 26, 2010
Palaeoporn 18

Look, I'm sorry about this. It's quite embarrassing, I know. But in my defense, I have blurred the naked Dickinsonia at the top right of the slab so as not to offend any sensibilities.
Anyhow, on to the real subject of this Palaeoporn, the structure with the highly descriptive and emotive name - "Form D". Yeah . . . "Form D" . . . no, honestly!
The Form D is the . . . um . . . rather flaccid structure above the compass. Form D was the name given to a group of thick, long, trace fossils of unknown origin. While they are not common, it doesn't take much poking about to expose them.
Given the lack of structure it was hard to attribute a source for the traces (they are not ribbed for example) thus making it difficult to erect a reasonable phylogeny and nomenclature. So they got stuck with the name "Form D" - as they were the fourth type of trace structures to be catagorised. Recently, they are thought to be, not trace fossils, but body parts. They appear to be association with the form Phyllozoon.
 Phyllozoon Hanseni (yellow arrows) and Form D (blue arrows)
Phyllozoon Hanseni (yellow arrows) and Form D (blue arrows)from the Ediacaran of Canada.
Phyllozoon is a more-or-less bisymmetrical leaf-shaped structure composed of a series of 'tubes' which meet at a central zigzag suture. There seems to have been only one layer of tubes forming the 'vanes' of the frond, with a stiff outer covering. Numerous individuals are commonly scattered over bedding planes, sometimes overlapping. The fronds have obvious polarity (top and bottom) and their common association with Form D is too frequent to be chance. It is possible that Form D is actually the stolon to which the Phyllozoon fronds were attached. They may well have looked something like this below, except Phyllozoon probably wasn't green.

I suppose we should be grateful that Phyllozoon wasn't called Phallozoon . . .
Sunday, September 19, 2010
Proterozoic Sponges Claim Doesn't Hold Water
The geologic account of ancient life is plagued with reports that do not withstand critical assessment. This is a special problem in the older rocks, where reports of spurious records continue to dilute the authentic record of evolution on the primitive Earth . . .
. . . It is left to the reader to draw conclusions about similar instances not here alluded to; the record of alleged pre-Phanerozoic life is full of them.
Preston Cloud (1973)
Harsh perhaps, but Preston Cloud's words should be engraved on the cover of every Proterozoic geologist's field notebook.
Frankly, the Proterozoic is weird. Most of what you think is organic, isn't. The vast majority of the rest is microbial mats. And the stuff you really, really think could be metazoan, is usually microbial mats playing silly buggers.
I don't want to become a party pooper (and no that's not a plug for Bora's pootopia). I don't, I really don't.
I want to be able to yell and scream about some new find that pushes our knowledge of the early evolution of metazoans back well into the Proterozoic. I do, I really do.
But I can't.
In Nature Geoscience, Maloof et. al discuss possible sponge-grade metazoans from the approx. 640 million year old Trezona Formation in the Flinders Ranges of South Australia.
The Trezona Formation just underlies the Marinoan glacial deposits (dated to approx. 635 million years ago) which mark the final throw of the of the Cryogenian dice, and are overlain by rocks of the Ediacaran Period. The Trezona marks the onset of the Marinoan glaciation and contains shallow water stromatolite flake breccia and bioclast packstones filling gaps between stromatolite heads:
. . . we identified a great diversity of bioclasts. Most packstones contain clasts of probable microbial origin, such as spalled flakes of adjacent stromatolite laminae and ripped-up and rolled-up sediments with cohesion enhanced by the presence of microbial mats. However, many bioclasts have anvil, wishbone, ring, and perforated slab morphologies that are difficult to assign to an abiotic roll-up or bacterial mat origin. in addition, the red colour and calcite composition of these distinctively shaped clasts are unique to the packstones (and even packstone clasts entrained in the overlying Elatina Fm diamictite as far as 65km from the nearest Trezona Fm stromatolite reef outcrop) and are not found in situ in the layers elsewhere in the Trezona Fm that could have been brecciated and transported. Therefore we suspect that the 1-cm-scale red bioclasts represent the remnants of a community of organisms endemic to the stromatolite-packstone environment. (Maloof et.al 2010.)This is what we are talking about

including "anvils" (d) and "perforated slabs" (g). (Maloof et.al 2010.)
2D thin sections show the clasts are composed of a mixture of opaque clays, quartz grains and micro-crystalline calcite, finer than the surrounding matrix, with a sharp, continuous contact with micritic rims

The authors then describe 3D shapes using serial sectioning (grid a bit off, photograph, grind a bit off, photograph). These 3D images show a three-dimensional network of 1mm diameter interconnected tubes. The tubes are also lined with micrite.
 Three-dimensional reconstruction of the Trezona Formation
Three-dimensional reconstruction of the Trezona Formationstructures. (Maloof et.al 2010.)
So we have stromatolites with infilling packstones contain a diverse set of bioclasts representing chips from associated stromatolites and ripped up and rolled sediments that have been glued together by microbial mats. However, the paper claims that some of these bioclasts have shapes, include anvil, wishbone, ring and perforated slabs, that cannot be caused by nonbiological actions for the following reasons:
1) The shapes cannot be formed by rock chips
2) The red colour and micrite composition is unique to the deposits - no source of the chips can be found.
3) The micrite coating in the tubes and around the clasts may represent weakly calcified cell layers.
4) The interconnected 3D tube structures and the 3D symmetry support a sponge hypothesis
1 and 2
OK, lets split these up. The first two deal with the lithology and composition of the clasts. The authors say that the clast shapes produced cannot be formed from mud chips or bits of stromatolite, and that is true. It is also true that there are no lithologies similar in colour or composition to the clasts anywhere else in the Trezona Formation. This means that the shapes cannot represent chips eroded off earlier formed mudstones because there are nor similar lithologies or rock types in the area, and even if there were, rock chips cannot create the shapes seen. But there are problems with using that as supporting evidence for a biological origin. There are other explanations. Other abiogenic explanations.
The local explanation for the clasts is that they are mud flakes not mud chips, that is, not mud chips eroded off pre-existing rocks, but mud flakes formed when muds are deposited in ephemeral or short-lived deposits, such as ponds after flooding surrounding areas, or overbank deposits.

These muds settle and dry out and can form all sorts of shapes.

In some instances the mud flakes can completely roll up

{kind=link}
{kind=link}
Check out one of the rolled mud flakes pictured above compared with a couple of the clast shapes

Clearly some mud flakes can take on the shapes that mud chips cannot, and can form some of the shapes present in the Trezona Formation that are being used as evidence for a biologic origin.
 But it gets worse. See the arrowed clasts in the image with the "anvil" shape (at right). These are clearly mud flakes. They show a common curled profile that can be seen in the examples of recent drying muds above. Nor do they contain the tubes that are supposed to be a feature of the "sponge" clasts.
But it gets worse. See the arrowed clasts in the image with the "anvil" shape (at right). These are clearly mud flakes. They show a common curled profile that can be seen in the examples of recent drying muds above. Nor do they contain the tubes that are supposed to be a feature of the "sponge" clasts.Now, here's the interesting bit. Both the obvious mud flakes and the "sponge" clasts are the same colour AND have the same composition (opaque clays and microcrystalline calcite, with the occasional very fine quarts grain - see photo-micrographs above) in other words your typical mud overbank deposit. Especially since the Trezona is pretty much a lowstand deposit (lower sea level). Some of the limestones deposited prior to the Trezona Formation, when sea levels were higher, are now exposed, providing a source for carbonate and putting some distance between the Trezona depositional environment and any non-carbonate source material (meaning that only very fine grained material is likely to reach the depocentre).
I think it is clear that the red clasts are mud flakes from overbank deposits, rolled up and redeposited between the stromatolites. In other words, waters carrying red carbonate mud, overtop riverbanks and settle into ponds. The mud settle out and the ponds dry up. The mud then dries, with the carbonate providing a good cement. The mud cracks, curls and eventually are dispersed by winds rain or floods into the nearby sea. The mud could be sourced from further inland on the Gawler Craton, and the carbonate sourced from limestones deposited during the period before the current regressive cycle, of which the Trezona Formation represents a lowstand or low sea level phase. The ephemoral nature of these overbank deposits, and their lack of lateral extent means that they do not contribute to the rock record, but dry out, crack, curl and are dispersed by wind and/or water. This also explains the occasional presence of "silica blebs" within the clasts. These represent very fine sand grains incorporated into the muds as they are deposited.
Clearly the mud flakes and "sponge" clasts have the same source - which is not organic. The shapes and colour cannot be used as supporting evidence of an organic origin for the clasts.
The authors suggest that the red matrix could be a replacement:
Alternatively, the original organic skeletons could have been coated in a bacterial extracellular polymeric substance following their death. The chemical composition of the Fe, Na, K clays found in the Trezona Fm bioclasts is similar to that of augenic minerals precipitated by microbial biofilms during the replacement of soft tissues. The extracellular polymeric substance would have also formed a template for abiotic calcium precipitation. (Maloof et.al 2010.)Now if I'm reading that right, it looks like the authors are suggesting that the original skeleton of the "sponges" has been replaced by the red mud, since there is no evidence for spicules or any other type of a primary skeleton.
There are two problems with this explanation for the absence of a primary skeleton. The first is the fact that the red matrix composition is shared with clasts that are clearly mud flakes. The second problem goes to number three in their list of supporting evidence, that the micrite coating in the tubes and around the clasts may represent weakly calcified cell layers.
3
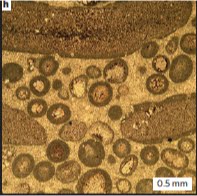 The authors comment that the "sponge" clasts are surrounded by a micrite rim (the dark outer rim around the clasts in the photo-micrograph at right. I don't know why they don't use the standard terminology - micrite envelope), which is also present lining the tubes. They suggest that:
The authors comment that the "sponge" clasts are surrounded by a micrite rim (the dark outer rim around the clasts in the photo-micrograph at right. I don't know why they don't use the standard terminology - micrite envelope), which is also present lining the tubes. They suggest that:the micrite of uniform thickness and texture coating both the exterior surface and interior canal walls could represent weakly clacified cell layers sandwiching the mesohyl of a sponge grade organism. (Maloof et.al 2010.)To their credit the authors then demolish this argument by pointing out that:
However, texturally similar (but less uniform thickness) micrite also coats peloids that do not seam to be part of the Trezona Fm organisms.Yes, that's a bit of a problem. It's a bit difficult to suggest that the micrite envelopes are evidence of mesohyl when almost everything in the deposit has them.
Micrite envelopes are well known and are generally considered to be formed from filamentous organisms (bacterial, algal or fungal) who's filaments calicify and coalesce into a intertwined mesh around the clasts. In this case the areas between stromatolites are colonised by numerous filamentous organisms that grow around all the clasts present and eventually form the micritic envelopes. So having micritic envelopes around the "sponge" clasts and within the tubes cannot be used as evidence of sponge-grade tissues.
But there's another problem.
As stated above, the authors suggest that the mud matrix could be a replacement for the original skeleton structure around the tubes. The presence of the micrite envelopes is really problematic here. The authors comment that:
However, the contact between micrite coating and mixed clay-chert-calcite interiors is usually sharp, with no evidence of diffusive or porosity-following micritization. (Maloof et.al 2010.)This means that any replacement of the original skeleton must have occurred prior to the micrite envelope being laid down. However, the tubes must have been sealed during replacement as there is no replacement material in them. After replacement, the tubes must have opened again to allow the micrite envelope to form.
Unlikely.
The evidence suggests that the red matrix is not a replacement, but a primary feature.
4
Which brings us to number 4, that the interconnected 3D tube structures and the 3D symmetry support a sponge hypothesis. I'll say up front that I do not know what the tubes are. They could be sponge tubes, but the evidence for that is equivocal at best.
A more likely explanation is that the "sponge" clasts are mud flakes that have been coated by filamentous organisms, rolled around and glued together. The tubes would then be where the filamentous outer coating of individual clasts, acting as a buffer and holding the clasts apart as a number of clasts were bound together.
It is interesting to note that an analysis of similar lithologies from rocks of the the equivalent Cryogenian interval south of Adelaide found;
. . . a light greyish microcrystalline limestone in which numerous flakes of grey calcareous mud are set. The flakes are typically intraformational and a gradation is noticed from intraformational breccia to edgewise conglomerate. Most of the flakes are flat, but there is a tendency to turn at the edges; length varies from several to 20 mm. in thin section or on polished faces there is a superficial resemblance to annelids (reference), but this is quite lost when the third dimension is considered. (Sprigg 1942)In the Proterozoic, weirdness rules. This makes interpretation difficult. I don't think the evidence presented in the paper is enough for a sponge interpretation to hold water. However, we've been misinterpreting the Proterozoic for some time, and the authors are in good company.
Cloud, P. (1973) Pseudofossils: A Plea for Caution. Geology, v. 1, p. 123-127.
Maloof, A., Rose, C., Beach, R., Samuels, B., Calmet, C., Erwin, D., Poirier, G., Yao, N., & Simons, F. (2010). Possible animal-body fossils in pre-Marinoan limestones from South Australia Nature Geoscience, 3 (9), 653-659 DOI: 10.1038/NGEO934
Sprigg, R.C. (1942) The Geology of the Eden-Moana Fault Block. Transactions of the Royal Society of South Australia, 66(2), 184-214. Download
Thursday, September 9, 2010
Please Give to DonorsChoose

OK I'm back and finalising a couple of posts. In the meantime DonorsChoose is running again.
DonorsChoose.org is an online charity that makes it easy to help US school students in need. (Yes I'm Australian, but this is important.)
American public school teachers post classroom project requests on DonorsChoose.org. Requests are for arts, science, music, language, civics, sports, etc.
You can browse project requests and give any amount to the one that inspires you. Once a project reaches its funding goal, the materials are sent to the school.
Education is important. If you have a few dollars to spare please consider supporting teachers trying to give their students a good education.
A number of the projects are close to being fully funded, a few dollars could make all the difference.
Wednesday, September 1, 2010
We've Lost Cedric
 Cedric the Tasmanian Devil has been put down.
Cedric the Tasmanian Devil has been put down.Cedric was at the forefront of research into the deadly Devil Facial Tumor Disease, which at last estimate was infecting 60 per cent of the wild devil population in Tasmania.
He made waves in 2007 when he was injected with a strain of DFTD and survived. He even survived a second strain with only a couple of minor tumours, which were removed.
Cedric had shown an immune response to the disease, and was otherwise healthy. It was hoped that he would lead researchers to a treatment or vaccine for the disease, but an X-ray last week showed a number of tumours in the lungs, and it was decided to euthanase the 6-year-old Cedric.
Damn.
Donations to help the Tassie Devils can be made here.
Tuesday, August 31, 2010
Cutticcini's Profile Enlargement
The wingnut Virginian Attorney General Ken Cuccinelli has had a setback in his witchhunt of Professor Michael Mann. Cuccinelli was asking for all documents from the University of Virginia concerning professor Mann's research while at UVa, to "investigate" any possible fraud committed by Professor Mann in his grants received to study climate change. A judge has ruled that Cuccinelli failed to show a sufficient “reason to believe” that UVa possessed any documents related to Mann that suggested a fraud occurred. Cuccinelli will continue harrassing UVa and Mann on the taxpayer's dime.
Cuccinelli is a climate change denier, who has also directed all state schools and colleges to remove sexual orientation references from their non-discrimination policies (in other words removed discrimination protection for gays and lesbians), and sued over the constitutionality of federal health care reform. All on the taxpayer's dime.
Cuccinelli is doing nothing more that using taxpayer money to fund a major profile enlargement, which is odd given his opposition to publicly funded healthcare.
Cuccinelli is a climate change denier, who has also directed all state schools and colleges to remove sexual orientation references from their non-discrimination policies (in other words removed discrimination protection for gays and lesbians), and sued over the constitutionality of federal health care reform. All on the taxpayer's dime.
Cuccinelli is doing nothing more that using taxpayer money to fund a major profile enlargement, which is odd given his opposition to publicly funded healthcare.
Sunday, August 15, 2010
Evolution Among the Trilobites Part 1
The last Palaeoporn featured the growth stages of meraspid, or juvenile, Estaingia. This post will use those meraspids and a bunch of holaspids, or adults, to measure the growth patterns of Estaingia, or it’s ontogeny.
Why? Because it’s important. It allows us to plot the growth patterns and if you know the growth patterns you can used them to look at evolutionary relationships.
If you want to look at evolutionary relationships between species there is one, and only one rule.
Ya gotta know what the kids are doing.
If you don’t know what the kids are doing, you can get, as some vertebrate palaeontologists are now finding, evolutionarily embarrassed. Ontogeny can provide clues to evolutionary relationships.
Studies of ontology requires the measurement of various features to show how they change with time or size (usually the two are interchangeable – in trilobites anyway). The changes in one feature are then compared with changes in another.
 At right is the head of an average trilobite showing which bits are which. This post will be concerned only with the head and what happens to it during ontogeny. Actually its even more restricted than just the head. We'll only be looking at the cranidium. That's the central area including the bulbous glabella. Why? well we need to take a bunch of measurements over a range of specimens to build up a picture of the changes that occur during ontogeny. The trouble with the librigene or free cheeks is that they are, well . . . free, and tend to be lost (as pointed out in Palaeoporn 14 and 16). The cranidium is usually left behind and so is a better subject for measurements.
At right is the head of an average trilobite showing which bits are which. This post will be concerned only with the head and what happens to it during ontogeny. Actually its even more restricted than just the head. We'll only be looking at the cranidium. That's the central area including the bulbous glabella. Why? well we need to take a bunch of measurements over a range of specimens to build up a picture of the changes that occur during ontogeny. The trouble with the librigene or free cheeks is that they are, well . . . free, and tend to be lost (as pointed out in Palaeoporn 14 and 16). The cranidium is usually left behind and so is a better subject for measurements.
.jpeg) At right is a cranidium of an Estaingia. The relevant measurements that we are interested in are marked. CW is cranidial width, which is literally the distance between the eyes. PGF is Pre-Glabella Field which is the area in front of the large bulbous glabella. (Incidentally the pre-glabella field only runs to the shallow trench towards the front of the cranidium. In front of the trench is the doublure. That is folded underneath the cranidium in life, along the trench, and pops up during moulting (see Palaeoporn 16). GL is the glabella length. Adding PGF and GL gives us a value for the length of the craniduum. POG is Pre-Orbital Glabella which is the bit of the glabella that lies in front of a line drawn between the front tip of the eyes. AF-EL is the distance between the end of the Axial Furrow (the trench behind the glabella and the back tip of the Eye Lobe.
At right is a cranidium of an Estaingia. The relevant measurements that we are interested in are marked. CW is cranidial width, which is literally the distance between the eyes. PGF is Pre-Glabella Field which is the area in front of the large bulbous glabella. (Incidentally the pre-glabella field only runs to the shallow trench towards the front of the cranidium. In front of the trench is the doublure. That is folded underneath the cranidium in life, along the trench, and pops up during moulting (see Palaeoporn 16). GL is the glabella length. Adding PGF and GL gives us a value for the length of the craniduum. POG is Pre-Orbital Glabella which is the bit of the glabella that lies in front of a line drawn between the front tip of the eyes. AF-EL is the distance between the end of the Axial Furrow (the trench behind the glabella and the back tip of the Eye Lobe.
Lets show some measurements - the length of the central portion of the trilobite head, the cranidium, compared with the width of the cranidium, and the length of the glabella, for each specimen, as seen below
.jpeg) Here the measurements fall on a straight line. This is called isometric growth, where the ratio of the two features does not vary with growth. In example A, the ratio of the length of the cranidium to the width of the cranidium does not change with growth. In example B the ratio of the cradidial length to glabella length does not change. So during isotmetric growth there is no change in the shape. (What? Yes, OK, example A is not strictly isometric, since isometric growth always passes through the origin and a line drawn through the plot in A doesn’t. Just. But trust me, compared with what’s coming up next it’s pretty much isometric.)
Here the measurements fall on a straight line. This is called isometric growth, where the ratio of the two features does not vary with growth. In example A, the ratio of the length of the cranidium to the width of the cranidium does not change with growth. In example B the ratio of the cradidial length to glabella length does not change. So during isotmetric growth there is no change in the shape. (What? Yes, OK, example A is not strictly isometric, since isometric growth always passes through the origin and a line drawn through the plot in A doesn’t. Just. But trust me, compared with what’s coming up next it’s pretty much isometric.)
Isometric growth can be very useful in confirming that specimens belong to a particular species. In the examples above, the red dots represent meraspid Estaingia and the blue dots holaspid, or adult forms. The fact that they all line up on a straight line is supporting evidence that the meraspids do belong to Estaingia. This is important because species are defined by a “type” specimen which is normally an adult, and so it can be quite difficult to place meraspids in the correct species if the growth patterns vary considerably from the adult.
Oh yes, the black dots at the far end of both graph trends represents a holaspid Estaingia from a different location and a slightly younger deposit, at Cymbric Vale in New South Wales (NSW Estaingia). Thought to be the same species as E. bilobata discussed here from the Emu Bay Shale, but also though not to be for a couple of reasons. It'll feature later but see how it plots along the same trend as the other specimens? good evidence that it is E. bilobata, but I'm getting ahead of myself.
Frankly, however, isometric growth while it has it’s place, is a bit boring. Meraspid 1 looks pretty much like the adult form. (no problem in putting the meraspids in the correct species there then.)
Just as well then that not all growth is isometric. In fact most growth patterns are not isometric, but anisometric. Anisometric growth, as you may suspect, is where the ratio does not remain constant, but changes during growth, and so shape changes during growth. Which means the meraspid can look distinctly different from the holaspid.

Holy rotating vectors Batman!
Now it gets interesting. There is only so much you can do with straight lines. Everything is better with curves! Anisometric growth changes are not random. They don’t occur because the merasid didn’t like the look of itself in the mirror. The represent hereditary traits. In the two graphs above, A shows Cranidial Width against Axial Furrow to posterior of Eye Lobe (as a percentage of cranidial lenght) (CW/AF-EL) and B shows Cranidial Length against Pre-Glabella Field (as a percentage of cranidial length) (PGF/GL+PGF).
Here the difference in growth patterns between the merasipd and holaspids is marked. This is telling us important information about how the final look of the holaspid form is achieved, and what pattern of growth occurred.
In graph A the feature being measured is the curvature of the eye. In early meraspids, the eye curvature is quite shallow and so the lower end of the eye is a significant distance from the Axial Furrow. As the meraspid grows, the eye curvature becomes more pronounced and the distance between the lower end of the eye and the axial furrow decreases. At the end of the meraspid stage, the distance (as a percentage of cranidial length) is around 0.25. In the holaspid stage this trend ceases, the eye reaches its maximum curvature, and doesn't change again, so the value remains at around 0.25. The NSW Estaingia plots along this holaspid trend albeit at a larger size that those at Emu Bay, but again, it is good evidence that NSW Estaingia is Estaingia bilobata.
In Graph B the preglabella field as a percentage of total cranidial length is plotted against cranidial length. Here the trends are different. In the meraspids, the preglabella field increases with growth until the Holaspid stage is reached. Then the trend is reversed and the preglabella field begins to reduce in size with growth, albeit at a slower rate than the meraspid rate. The NSW Estaingia again plot on the holaspid trend for Estaingia bilobata providing more evidence that the NSW Estaingia is E. bilobata.
One last graph which shows the preorbital glabella or POG (the portion of the glabella in front of the eyes).
 Note that this is a mirror of the pregalbella field plot, and provides an explanation for the pregalbella field changes. The preorbital glabella in the meraspids decreases in size relative to cranidial length. In other words, as the cranidium increases in size, the preorbital glabella is increasing at a slower rate. This means that the area in front of it has to increase in size. So the increase in the pregalbella field we saw in the previous chart is a result of the preoccular glabella not keeping up during meraspid growth. However, in the holaspid stage, this trend is reversed and the preoccular glabella starts to grow at a more rapid pace than the cranidium generally, and begins to represent more and more of the total cranidial length. Thus the preglabella field begins to decrease as a proportion of the total cranidial length. Once again the NSW Estaingia plot among the Estaingia bilobata and along the trend. We can say with some confidence then that NSW Estaingia is a larger version of Estaingia bilobata.
Note that this is a mirror of the pregalbella field plot, and provides an explanation for the pregalbella field changes. The preorbital glabella in the meraspids decreases in size relative to cranidial length. In other words, as the cranidium increases in size, the preorbital glabella is increasing at a slower rate. This means that the area in front of it has to increase in size. So the increase in the pregalbella field we saw in the previous chart is a result of the preoccular glabella not keeping up during meraspid growth. However, in the holaspid stage, this trend is reversed and the preoccular glabella starts to grow at a more rapid pace than the cranidium generally, and begins to represent more and more of the total cranidial length. Thus the preglabella field begins to decrease as a proportion of the total cranidial length. Once again the NSW Estaingia plot among the Estaingia bilobata and along the trend. We can say with some confidence then that NSW Estaingia is a larger version of Estaingia bilobata.
Next time how these growth plots can provide information on evolutionary relationships.
Why? Because it’s important. It allows us to plot the growth patterns and if you know the growth patterns you can used them to look at evolutionary relationships.
If you want to look at evolutionary relationships between species there is one, and only one rule.
If you don’t know what the kids are doing, you can get, as some vertebrate palaeontologists are now finding, evolutionarily embarrassed. Ontogeny can provide clues to evolutionary relationships.
Studies of ontology requires the measurement of various features to show how they change with time or size (usually the two are interchangeable – in trilobites anyway). The changes in one feature are then compared with changes in another.
 At right is the head of an average trilobite showing which bits are which. This post will be concerned only with the head and what happens to it during ontogeny. Actually its even more restricted than just the head. We'll only be looking at the cranidium. That's the central area including the bulbous glabella. Why? well we need to take a bunch of measurements over a range of specimens to build up a picture of the changes that occur during ontogeny. The trouble with the librigene or free cheeks is that they are, well . . . free, and tend to be lost (as pointed out in Palaeoporn 14 and 16). The cranidium is usually left behind and so is a better subject for measurements.
At right is the head of an average trilobite showing which bits are which. This post will be concerned only with the head and what happens to it during ontogeny. Actually its even more restricted than just the head. We'll only be looking at the cranidium. That's the central area including the bulbous glabella. Why? well we need to take a bunch of measurements over a range of specimens to build up a picture of the changes that occur during ontogeny. The trouble with the librigene or free cheeks is that they are, well . . . free, and tend to be lost (as pointed out in Palaeoporn 14 and 16). The cranidium is usually left behind and so is a better subject for measurements..jpeg) At right is a cranidium of an Estaingia. The relevant measurements that we are interested in are marked. CW is cranidial width, which is literally the distance between the eyes. PGF is Pre-Glabella Field which is the area in front of the large bulbous glabella. (Incidentally the pre-glabella field only runs to the shallow trench towards the front of the cranidium. In front of the trench is the doublure. That is folded underneath the cranidium in life, along the trench, and pops up during moulting (see Palaeoporn 16). GL is the glabella length. Adding PGF and GL gives us a value for the length of the craniduum. POG is Pre-Orbital Glabella which is the bit of the glabella that lies in front of a line drawn between the front tip of the eyes. AF-EL is the distance between the end of the Axial Furrow (the trench behind the glabella and the back tip of the Eye Lobe.
At right is a cranidium of an Estaingia. The relevant measurements that we are interested in are marked. CW is cranidial width, which is literally the distance between the eyes. PGF is Pre-Glabella Field which is the area in front of the large bulbous glabella. (Incidentally the pre-glabella field only runs to the shallow trench towards the front of the cranidium. In front of the trench is the doublure. That is folded underneath the cranidium in life, along the trench, and pops up during moulting (see Palaeoporn 16). GL is the glabella length. Adding PGF and GL gives us a value for the length of the craniduum. POG is Pre-Orbital Glabella which is the bit of the glabella that lies in front of a line drawn between the front tip of the eyes. AF-EL is the distance between the end of the Axial Furrow (the trench behind the glabella and the back tip of the Eye Lobe.Lets show some measurements - the length of the central portion of the trilobite head, the cranidium, compared with the width of the cranidium, and the length of the glabella, for each specimen, as seen below
.jpeg) Here the measurements fall on a straight line. This is called isometric growth, where the ratio of the two features does not vary with growth. In example A, the ratio of the length of the cranidium to the width of the cranidium does not change with growth. In example B the ratio of the cradidial length to glabella length does not change. So during isotmetric growth there is no change in the shape. (What? Yes, OK, example A is not strictly isometric, since isometric growth always passes through the origin and a line drawn through the plot in A doesn’t. Just. But trust me, compared with what’s coming up next it’s pretty much isometric.)
Here the measurements fall on a straight line. This is called isometric growth, where the ratio of the two features does not vary with growth. In example A, the ratio of the length of the cranidium to the width of the cranidium does not change with growth. In example B the ratio of the cradidial length to glabella length does not change. So during isotmetric growth there is no change in the shape. (What? Yes, OK, example A is not strictly isometric, since isometric growth always passes through the origin and a line drawn through the plot in A doesn’t. Just. But trust me, compared with what’s coming up next it’s pretty much isometric.)Isometric growth can be very useful in confirming that specimens belong to a particular species. In the examples above, the red dots represent meraspid Estaingia and the blue dots holaspid, or adult forms. The fact that they all line up on a straight line is supporting evidence that the meraspids do belong to Estaingia. This is important because species are defined by a “type” specimen which is normally an adult, and so it can be quite difficult to place meraspids in the correct species if the growth patterns vary considerably from the adult.
Oh yes, the black dots at the far end of both graph trends represents a holaspid Estaingia from a different location and a slightly younger deposit, at Cymbric Vale in New South Wales (NSW Estaingia). Thought to be the same species as E. bilobata discussed here from the Emu Bay Shale, but also though not to be for a couple of reasons. It'll feature later but see how it plots along the same trend as the other specimens? good evidence that it is E. bilobata, but I'm getting ahead of myself.
Frankly, however, isometric growth while it has it’s place, is a bit boring. Meraspid 1 looks pretty much like the adult form. (no problem in putting the meraspids in the correct species there then.)
Just as well then that not all growth is isometric. In fact most growth patterns are not isometric, but anisometric. Anisometric growth, as you may suspect, is where the ratio does not remain constant, but changes during growth, and so shape changes during growth. Which means the meraspid can look distinctly different from the holaspid.

Now it gets interesting. There is only so much you can do with straight lines. Everything is better with curves! Anisometric growth changes are not random. They don’t occur because the merasid didn’t like the look of itself in the mirror. The represent hereditary traits. In the two graphs above, A shows Cranidial Width against Axial Furrow to posterior of Eye Lobe (as a percentage of cranidial lenght) (CW/AF-EL) and B shows Cranidial Length against Pre-Glabella Field (as a percentage of cranidial length) (PGF/GL+PGF).
Here the difference in growth patterns between the merasipd and holaspids is marked. This is telling us important information about how the final look of the holaspid form is achieved, and what pattern of growth occurred.
In graph A the feature being measured is the curvature of the eye. In early meraspids, the eye curvature is quite shallow and so the lower end of the eye is a significant distance from the Axial Furrow. As the meraspid grows, the eye curvature becomes more pronounced and the distance between the lower end of the eye and the axial furrow decreases. At the end of the meraspid stage, the distance (as a percentage of cranidial length) is around 0.25. In the holaspid stage this trend ceases, the eye reaches its maximum curvature, and doesn't change again, so the value remains at around 0.25. The NSW Estaingia plots along this holaspid trend albeit at a larger size that those at Emu Bay, but again, it is good evidence that NSW Estaingia is Estaingia bilobata.
In Graph B the preglabella field as a percentage of total cranidial length is plotted against cranidial length. Here the trends are different. In the meraspids, the preglabella field increases with growth until the Holaspid stage is reached. Then the trend is reversed and the preglabella field begins to reduce in size with growth, albeit at a slower rate than the meraspid rate. The NSW Estaingia again plot on the holaspid trend for Estaingia bilobata providing more evidence that the NSW Estaingia is E. bilobata.
One last graph which shows the preorbital glabella or POG (the portion of the glabella in front of the eyes).
 Note that this is a mirror of the pregalbella field plot, and provides an explanation for the pregalbella field changes. The preorbital glabella in the meraspids decreases in size relative to cranidial length. In other words, as the cranidium increases in size, the preorbital glabella is increasing at a slower rate. This means that the area in front of it has to increase in size. So the increase in the pregalbella field we saw in the previous chart is a result of the preoccular glabella not keeping up during meraspid growth. However, in the holaspid stage, this trend is reversed and the preoccular glabella starts to grow at a more rapid pace than the cranidium generally, and begins to represent more and more of the total cranidial length. Thus the preglabella field begins to decrease as a proportion of the total cranidial length. Once again the NSW Estaingia plot among the Estaingia bilobata and along the trend. We can say with some confidence then that NSW Estaingia is a larger version of Estaingia bilobata.
Note that this is a mirror of the pregalbella field plot, and provides an explanation for the pregalbella field changes. The preorbital glabella in the meraspids decreases in size relative to cranidial length. In other words, as the cranidium increases in size, the preorbital glabella is increasing at a slower rate. This means that the area in front of it has to increase in size. So the increase in the pregalbella field we saw in the previous chart is a result of the preoccular glabella not keeping up during meraspid growth. However, in the holaspid stage, this trend is reversed and the preoccular glabella starts to grow at a more rapid pace than the cranidium generally, and begins to represent more and more of the total cranidial length. Thus the preglabella field begins to decrease as a proportion of the total cranidial length. Once again the NSW Estaingia plot among the Estaingia bilobata and along the trend. We can say with some confidence then that NSW Estaingia is a larger version of Estaingia bilobata.Next time how these growth plots can provide information on evolutionary relationships.
Subscribe to:
Comments (Atom)